Анатомия корня растений
Корень — фундаментальный орган высших растений, без которого невозможно представить их существование на суше. Он не только прочно фиксирует растение в почве, но и служит главным каналом поступления воды и минеральных веществ. Однако за кажущейся простотой скрывается сложная анатомическая организация, эволюционная история и поразительное разнообразие форм, позволяющее растениям адаптироваться к самым разным условиям.
Происхождение и отличие от побега
Корень возник в результате морфологической дифференциации осевых органов — теломов — древнейших высших растений, осваивавших сушу. Первыми структурами, предшествовавшими настоящему корню, стали подземные теломы-ризомоиды, поверхность которых покрывали ризоиды. Именно эти органы обеспечили первичную возможность поглощать воду и минеральные вещества из почвы. Позднее, предположительно после появления побега, в эволюционной последовательности возник полноценный корень.
Осевое происхождение корня подтверждается отсутствием существенных анатомических различий между ним и стеблем у некоторых высших споровых растений на ранних стадиях развития. У современных растений эти различия сводятся к деталям: и корень, и стебель обладают радиальной симметрией, апикальным ростом за счёт меристемы и участвуют в транспорте веществ. Однако корень характеризуется уникальным набором признаков: отсутствием листьев, наличием чехлика, покрывающего апикальную меристему, корневыми волосками для извлечения почвенного раствора, а также эндогенным ветвлением. В отличие от стебля, где боковые побеги закладываются экзогенно наружными частями конуса нарастания, у корня зачатки боковых корней формируются эндогенно — в глубине внутренних тканей, удалённых от апекса. Такое внутреннее закладывание обусловливает эффективное исследование объёма почвы и облегчает всасывание питательных веществ.
Не все высшие растения сохранили корни. Моховидные поглощают воду ризоидами. Псилотовидные (род Psilotum и Tmesipteris) вместо настоящих корней развивают корневищеподобные органы с дихотомически разветвлёнными образованиями, усыпанными ризоидами; эти структуры морфологически напоминают ризомоиды палеозойских риниевых, что подчёркивает их происхождение из древних подземных теломов. Кроме первично бескорневых, существуют и «вторично» бескорневые растения, утратившие корни при переходе к водному образу жизни (например, Salvinia из папоротниковидных, пузырчатка и роголистник из цветковых) или к гетеротрофному питанию (например, орхидея Laddia в болотистых районах северного полушария).
Функции корня в жизни растения
Корень служит основной опорной системой, фиксируя растение в субстрате. Главный корень проявляет положительный геотропизм и растёт вертикально вниз, обеспечивая прочную фиксацию ствола и листьев. Его кончик имеет конусовидную форму, плотную и прочную структуру, что позволяет преодолевать сопротивление и трение частиц почвы. В этом процессе важную роль играют корневые волоски: они не только поглощают воду, но и создают опору кончику, поддерживая его движение между твёрдыми частицами и усиливая закрепление.
Поглощающая функция обеспечивает водоснабжение и снабжение минеральными веществами. Через транспортную сеть корень доставляет растворы в вышерасположенные органы, а также участвует в синтезе аминокислот, нуклеотидов, алкалоидов, гормонов роста и других физиологически активных соединений. Одновременно корень выделяет ненужные метаболиты; клетки, отделившиеся от чехлика, уносят с собой балластные вещества, освобождая растение от их накопления.
Накопительная функция не конфликтует с основной: запасные вещества откладываются в корневой ткани, создавая энергетический резерв для неблагоприятных периодов. Кроме того, корень активно взаимодействует с микробным сообществом в ризосфере — тонком слое почвы толщиной 2–3 мм, прилегающем к корню. Здесь многочисленные микроорганизмы трансформируют трудноусваиваемые соединения в легкоусваиваемые формы и синтезируют биологически активные вещества, улучшая питание растения.
Сосущие корни образуют мутуалистические симбиозы с грибами — микоризу. Гифы микоризных грибов могут оплетать корень снаружи (эктотрофная микориза) или проникать в клетки, образуя внутри ветвящиеся клубки (эндотрофная микориза). В ряде случаев наблюдается сочетание обоих типов, что усиливает эффективность обмена питательными веществами и водой. Симбиотические отношения с азотфиксирующими бактериями также улучшают питание и водообеспечение, расширяя спектр доступных элементов. Корневые волоски в зоне поглощения существенно увеличивают всасывающую поверхность, завершая комплексную роль корня как структурного, питательного, синтетического и детоксикационного органа.
Типы корней и строение корневых систем
Корневая система растений может быть мономорфной, состоящей из корней одного типа, и гоморизной, образованной придаточными корнями. Гоморизная система характерна для споровых архегониальных растений, где зачатки придаточных корней формируются вблизи апексов побегов. У семенных растений придаточные корни закладываются дальше от листовых точек, а их количество определяется внешними условиями и механическими повреждениями. Главный корень развивается из зародышевого корешка и, проявляя положительный геотропизм, вертикально проникает в почву, образуя ось системы. От него отходят боковые корни, начинающие ветвиться выше зоны корневых волосков. Их развитие акропетальное: чем моложе боковой корень, тем ближе к кончику главного корня он расположен. Боковые корни не ориентированы по гравитации и растут горизонтально, формируя сетчатую структуру, из которой далее образуются новые боковые ветви.
Зачатки боковых корней располагаются на главном корне продольными рядами, известными как ортостихи. Их число варьирует у разных таксонов: у большинства растений их две, у гороха — три, у грецкого ореха, дуба и ольхи — шесть, у каштана — от восьми до четырнадцати, а у однодольных обычно ещё больше, хотя и не всегда стабильно. У тыквенных, молочайных и вьюнковых ортостихи находятся на одинаковом интервале (по четыре), в то время как у гвоздичных, крестоцветных и зонтичных они сближены попарно. Эта разница тесно связана со строением проводящей системы главного корня.
Придаточные (адвентивные) корни могут образовываться на гипокотиле (например, у берёзы, недотроги, бальзамина) или непосредственно на стебле. Стеблевые придаточные корни в эволюционном плане предшествовали главному и встречаются у всех споровых архегониальных растений. У семенных их появление не привязано к листьям и зависит от факторов среды. В подземной части также может развиваться корневище — подземный стебель с узлами и междоузлиями, служащий резервуаром питательных веществ. От него отрастают как корни, так и листья, а из почек формируются новые побеги. Особым типом специализированных органов являются корневые шишки и корнеплоды — утолщения корня, адаптированные к накоплению запасных веществ и отличающиеся по структуре от стеблевых клубней.
Адаптации корней к среде обитания крайне разнообразны. Воздушные корни снабжены аэренхимой для газообмена в условиях недостатка кислорода. Ксерофиты либо формируют глубоко проникающие корни для доступа к подземным водам, либо создают разветвлённую поверхностную сеть для быстрого поглощения дождевой влаги; у них часто выражена мощная кутикула и редкие устьица. Гидрофиты обладают развитой аэренхимой и слабо развитыми корневыми волосками, получая основную часть веществ через листья и используя корни преимущественно для закрепления.
Первичное анатомическое строение
U-образные утолщения оболочек характерны для клеток экзодермы и эндодермы. В корневищах ландыша такие утолщения наблюдаются в клетках первичной коры, прилегающих к эндодерме. Средняя часть коры обычно многослойна, а её внутренние клетки у ряда видов способны делиться периклинально; их потомки увеличиваются к периферии, растягиваются в тангенциальном направлении и выстраиваются в радиальные ряды. У растений влажных местообитаний, а также у многих пальм в первичной коре формируются рексигенные воздухоносные полости; в этой ткани могут концентрироваться группы волокон, а отдельные клетки подвергаются склерификации.
Стела (центральный цилиндр) корня состоит из перицикла и сложного радиального проводящего пучка, где радиальные тяжи первичной флоэмы чередуются по кругу с радиальными тяжами первичной ксилемы. В корне развивается единственный тяж прокамбия. Первые элементы протофлоэмы закладываются экзархно, то есть в наружной части прокамбия, а между их группами размещаются элементы протоксилемы с кольчатыми и спиральными утолщениями оболочек.
Дальнейшее развитие проводящих тканей идёт центростремительно: от внутренних сторон уже сформированных элементов дифференцируются более широкие и просветные элементы метафлоэмы и метаксилемы. Тяжи ксилемы и флоэмы распределяются по окружности стелы равномерно; их количество равно, но число тяжей варьирует. По этому признаку корни классифицируют как монархные (по одному тяжу), диархные (по два), триархные, тетрархные и полиархные (много тяжей). Монархные корни крайне редки (некоторые виды ужовника из папоротниковидных), диархные наиболее распространены, а полиархные характерны преимущественно для однодольных. Общий вид центрального цилиндра на поперечном срезе зависит от расположения элементов метаксилемы; у лука, тыквы и других растений стела наиболее широко просветна, и самый молодой водопроводящий элемент метаксилемы обычно находится в её центре.
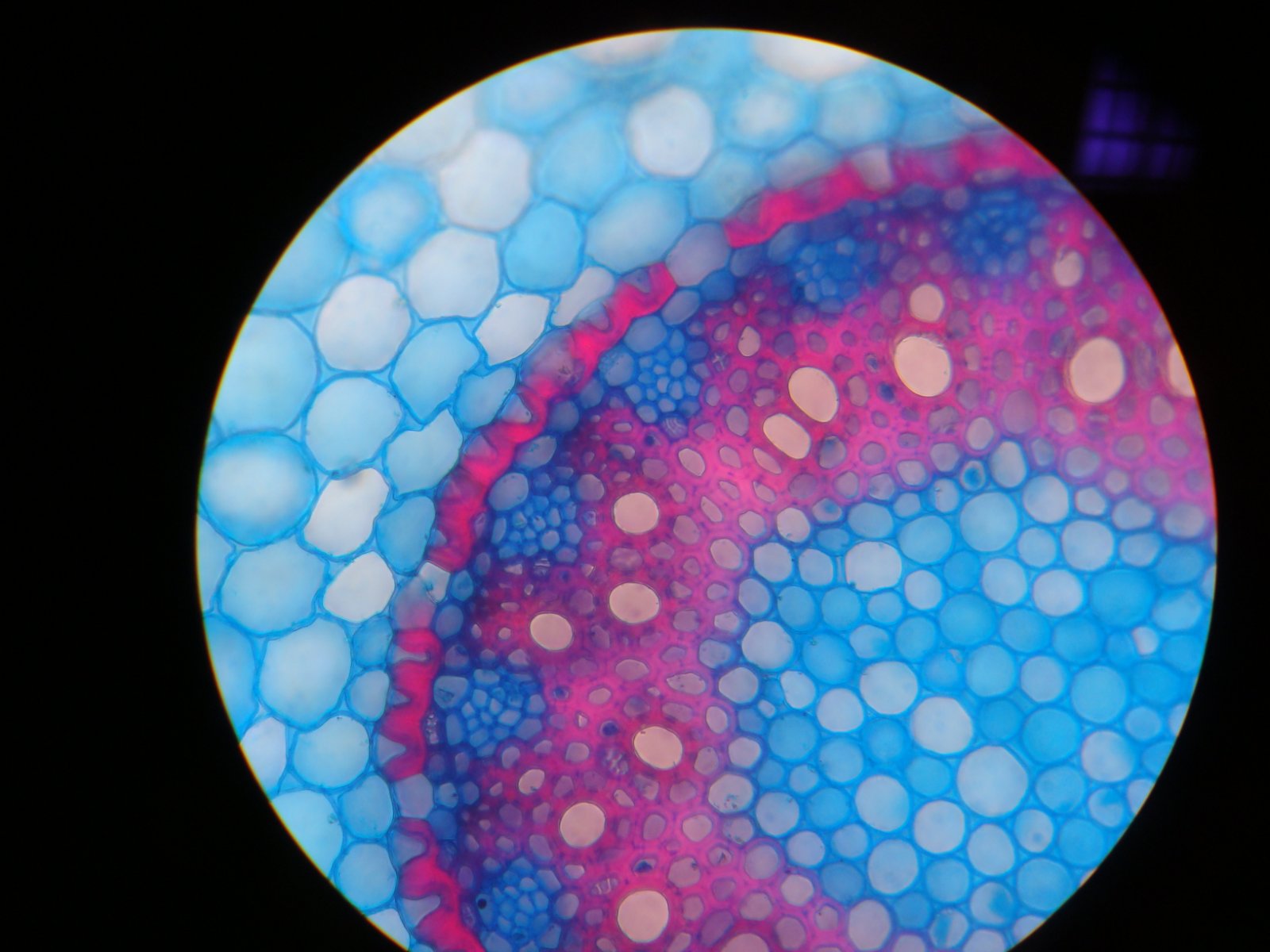
 Рис. 1. Схема первичного строения корня в поперечном сечении. Обозначены: стела (vscular cylinder), ксилема (xylem), флоэма (phloem), сердцевина (pith), прокамбий (procambium), перицикл (pericycle), эндодерма (endodermis), паренхима (parenchyma), ризодерма (epidermis), первичная кора (cortex). Стоит иметь ввиду, что ризодерма может не включаться в первичную кору.
Рис. 1. Схема первичного строения корня в поперечном сечении. Обозначены: стела (vscular cylinder), ксилема (xylem), флоэма (phloem), сердцевина (pith), прокамбий (procambium), перицикл (pericycle), эндодерма (endodermis), паренхима (parenchyma), ризодерма (epidermis), первичная кора (cortex). Стоит иметь ввиду, что ризодерма может не включаться в первичную кору.
Loki 66, CC BY-SA 3.0 http://creativecommons.org/licenses/by-sa/3.0/, via Wikimedia Commons
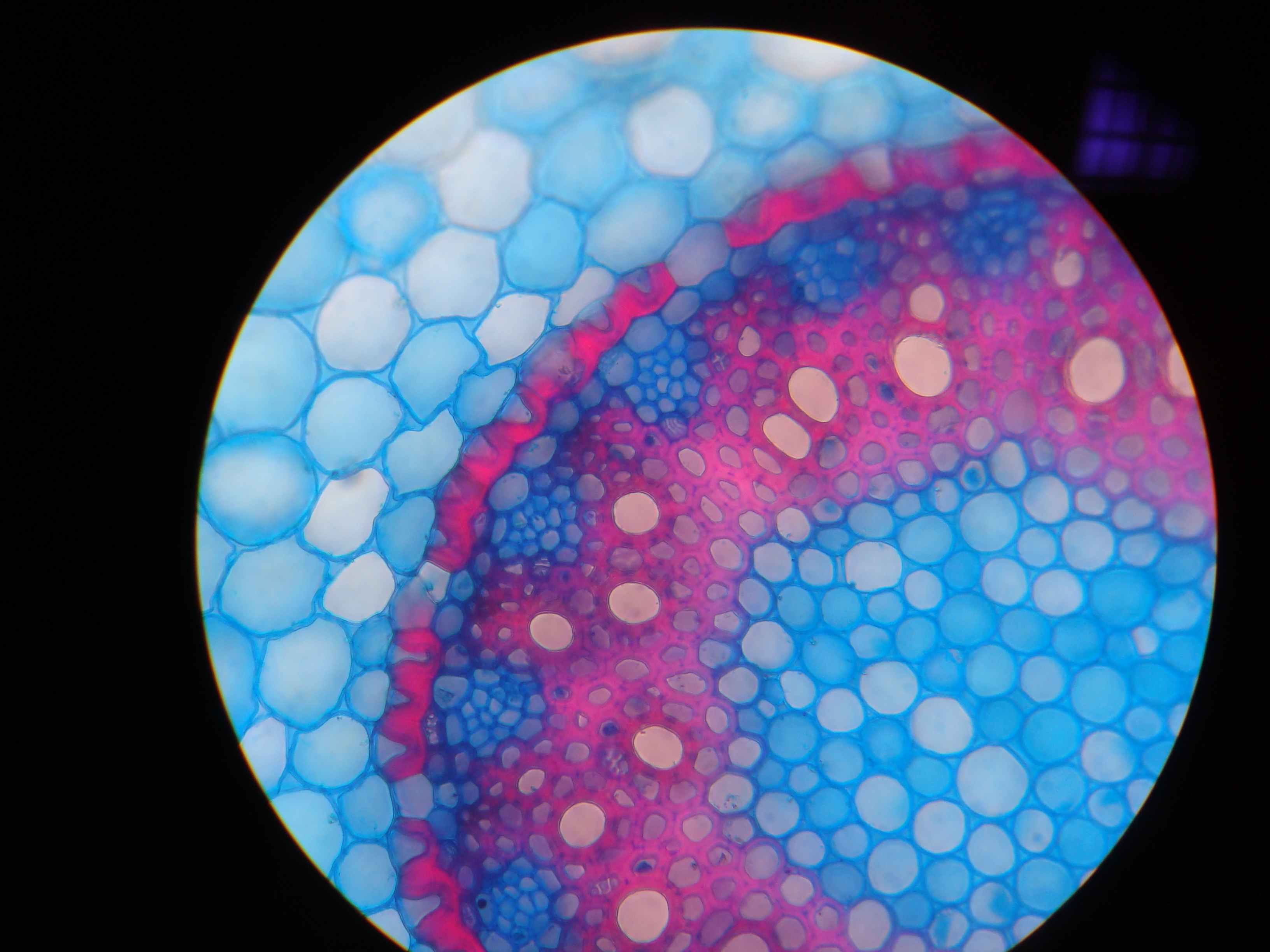 Рис. 2. Поперечный срез корня иглицы (Ruscus hypophyllum). Видны паренхима первичной коры, U-образные утолщения клеточных стенок эндодермы, перицикл, флоэма, ксилема, паренхима сердцевины.
Рис. 2. Поперечный срез корня иглицы (Ruscus hypophyllum). Видны паренхима первичной коры, U-образные утолщения клеточных стенок эндодермы, перицикл, флоэма, ксилема, паренхима сердцевины.
Вторичное утолщение и атипичный рост
При вторичном утолщении перицикл образует извилистый слой латеральной меристемы. Клетки, окружающие флоэмные тяжи, формируют пучковый камбий, тогда как части, охватывающие радиальные тяжи первичной ксилемы, образуют межпучковый камбий. Пучковый камбий начинает делиться ещё до формирования сплошного слоя, создавая элементы вторичной ксилемы внутрь и вторичной флоэмы наружу. Вторичная ксилема всегда объёмнее, поэтому она оттесняет камбий наружу, и первоначально вогнутая дуга камбия выпрямляется, приобретая выпуклую форму.
Деятельность пучкового камбия приводит к образованию наружных проводящих пучков, где самый внутренний элемент — вторичная ксилема, а самый наружный — остатки первичной флоэмы. Межпучковый камбий продуцирует лишь паренхимные клетки; их радиальные слои, отходя от тяжей первичной ксилемы, разделяют проводящие пучки, получая название паренхимных лучей (сердцевина в корне отсутствует).
В природе встречается несколько вариантов атипичного вторичного утолщения: неравномерная активность камбия, необычное расположение древесины и луба, деятельность нескольких камбиев одновременно или последовательно, а также утолщение с помощью особой меристематической зоны при полном отсутствии типичного камбия. Примером служит эксцентрический прирост древесины (эпиксилия) при наклонном или горизонтальном положении ветвей. На верхней (тяговой) стороне ветви древесина шире, а на нижней (креневой) — уже, причём креневая древесина богаче сосудами и беднее либриформом. Гипоксилия — иной тип: приросты древесины, богатые либриформом, толще на нижней стороне ветви (характерно для яблони, груши, облепихи). У чернодревесинных видов (акация, гледичия, софора) при эксцентричном росте луба волокна на нижней стороне ветви крупнее, с сильно одревесневшими оболочками, а на верхней — мельче, но с очень толстыми стенками.
В корнеплодах различия между вторичными тканями выражены ярко. У моркови вторичной ксилемы больше, чем флоэмы; во флоэме встречаются схизогенные вместилища с эфирными маслами. У редьки соотношение обратное, обе ткани богаты сочной паренхимой, а одревесневшие элементы редки; здесь присутствуют одиночные идиобласты. В корне свеклы после формирования двух коллатеральных пучков и паренхимных лучей основной камбий прекращает деятельность. Дальнейшее утолщение обеспечивается добавочными камбиями: первый закладывается в перицикл, и его клетки делятся периклинально. Внешний слой делений переходит в феллоген, внутренний остаётся камбием и формирует меристематическую ткань. Из неё возникает второй добавочный камбий, а внутренний слой образует коллатеральные пучки и межпучковую паренхиму.
Корневые шишки — это видоизменения боковых и придаточных корней, где запасы питательных веществ концентрируются в утолщённых участках. Утолщаться может апикальная часть корня (георгин, спаржа), средняя (зопник клубненосный) или образовываться четковидные вздутия (заячья капуста). Размеры шишек сильно варьируют: у ямса они могут достигать 40–50 кг, хотя чаще встречаются массой 5–6 кг, богатые крахмалом (в иностранной литературе их называют корневыми клубнями).
Вторичные ткани, формирующиеся при утолщении стебля и корня, включают производные камбия (вторичная ксилема, вторичная флоэма), феллогена (пробка, феллодерма, чечевички) и различные типы идиобластов. Идиобласты — структуры, отличающиеся по строению и функции от окружающих тканей; встречаясь внутри специализированных тканей, они утрачивают способность к делению и часто проявляются как склереиды в ассимилирующей ткани или эфирные вместилища. Вторичные ткани характерны для древесных растений и отсутствуют у мхов, современных хвощей, плаунов, папоротников (за редким исключением) и большинства однодольных покрытосеменных.
Заключение
Анатомия корня демонстрирует удивительное сочетание эволюционной консервативности и адаптивной пластичности. Базовый план строения, унаследованный от древних теломов, проявляется в радиальной симметрии, эндогенном ветвлении и тонкой структуре центрального цилиндра. В то же время, многообразие вторичных утолщений, образование специализированных запасающих органов, развитие микоризы и модификации корневых систем отражают тонкую настройку растения на конкретные условия среды — будь то засушливая пустыня, переувлажнённый субстрат или бедная питательными веществами почва. Именно корень, скрытый от глаз, во многом определяет успех и жизнестойкость растения, работая как якорь, насос и биохимическая лаборатория одновременно.