Анатомия стебля растений
Стебель — это не просто опора, связывающая корень и крону. Это сложная транспортная магистраль, живая архитектурная конструкция, способная адаптироваться к экстремальным нагрузкам, нехватке света и избытку влаги. Понимание анатомии стебля позволяет заглянуть в самую суть того, как растения эволюционировали, выживают и доминируют в самых разных экологических нишах. От микроскопических сосудов до мощных стволов — строение стебля отражает миллионы лет природной инженерии.
Наружные ткани и первичная кора
Эпидерма стеблей большинства высших растений имеет типовое строение: наружный слой клеток покрыт кутикулой, а в его пределах развиваются устьица, обеспечивающие газообмен. У ряда видов эпидерма дополнительно образует разнообразные волоски, которые усиливают защитные функции и участвуют в регуляции микроклимата поверхности.
Первичная кора у представителей семейства гвоздичных, многих лютиковых и ряда иных таксонов состоит из паренхимы, 1–2 наружных слоя которой обычно дифференцируются в хлоренхиму, способную к фотосинтезу. У большинства двудольных первичная кора упрощена и включает лишь два типа тканей — паренхиму и колленхиму. В некоторых стеблях, например у подсолнечника (Helianthus annuus) и кирказона (Centaurea cyanus), колленхима образует сплошной субэпидермальный слой, тогда как у зонтичных, губоцветных и подмаренника она присутствует в виде отдельных тяжей. Колленхимные клетки зачастую уголковые, однако в стеблях с интенсивным вторичным утолщением (как у подсолнечника) они могут принимать пластинчатую форму; рыхлая колленхима встречается у красавки (Centaurea cyanus).
Под слоем колленхимы располагается паренхима первичной коры, в которой часто встречаются зерна запасного крахмала, кристаллы оксалата кальция и дубильные вещества. Эта ткань богата межклетниками, и у некоторых растений (например, у подсолнечника и зонтичных) в ней образуются схизогенные вместилища для выделений. У молочая (Euphorbia) в первичной коре развиваются млечники и диффузно расположенные волокна склеренхимы. Самый внутренний паренхимный слой первичной коры называют крахмалоносным влагалищем; если его клеточные оболочки обладают пояском Каспари, этот слой считается эндодермой.
Стела и перицикл
Стела (центральный цилиндр) стебля состоит из перицикла, проводящего цилиндра и сердцевины. Перицикл, который может отсутствовать, обычно представляет собой паренхиму либо паренхиму в сочетании со склеренхимой, прилегающей к крахмалоносному влагалищу. Особенно выраженное развитие перицикла наблюдается у кирказона и многих гвоздичных, где он образует сплошные кольца, видимые на поперечных срезах. В некоторых случаях перицикл представлен отдельными тяжами слабо одревесневающей склеренхимы, что характерно для вьюнка (Convolvulus) и калистегии (Calystegia).
У растений, лишенных перицикла (сложноцветные, лютиковые, зонтичные и др.), крахмалоносное влагалище прилегает к тяжам толстостенных волокон, армирующих проводящие пучки. Эти волокна образуют протофлоэму и происходят из наружных клеток прокамбия, обеспечивая механическую поддержку и проводящую функцию взамен перицикла. У однодольных проводящие ткани располагаются в специфическом пространстве, но их особенности логично рассматривать в контексте общих стелярных элементов.
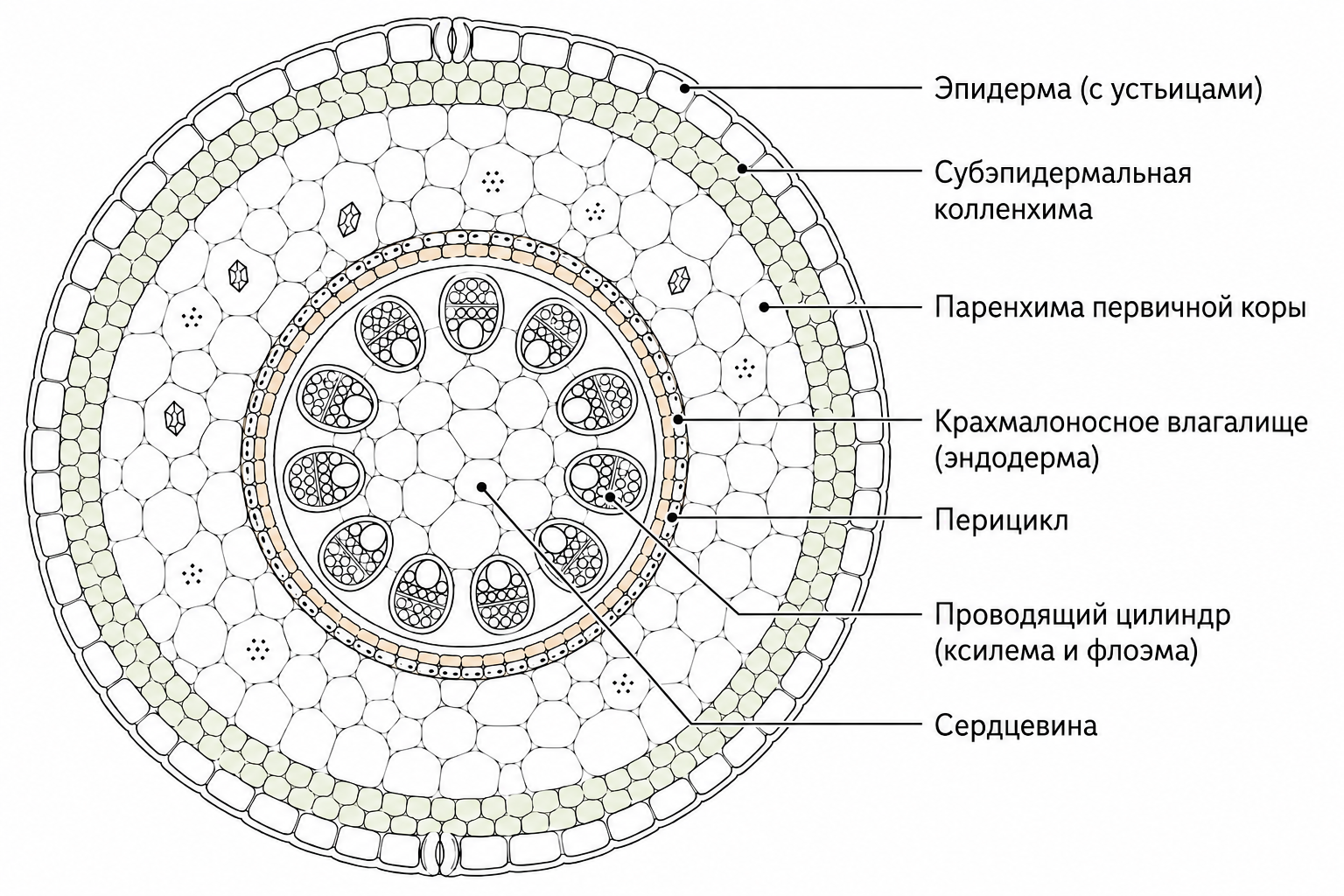 [ИЛЛЮСТРАЦИЯ: Схема поперечного среза стебля, показывающая расположение эпидермы, субэпидермальной колленхимы, паренхимы первичной коры, крахмалоносного влагалища, перицикла и проводящего цилиндра]
[ИЛЛЮСТРАЦИЯ: Схема поперечного среза стебля, показывающая расположение эпидермы, субэпидермальной колленхимы, паренхимы первичной коры, крахмалоносного влагалища, перицикла и проводящего цилиндра]
Меристемы и механизмы роста
Апикальная меристема стебля формируется в виде конуса нарастания, внутри которого выделяются три зоны. Фланговая зона расположена по бокам и отвечает за образование боковых органов; медуллярная зона образует паренхимные клетки будущей сердцевины, а основная меристема находится как в центральной части стебля, так и под протодермой на периферии. Из фланговой зоны формируется эвмеристема — полый цилиндр, слегка расширяющийся вниз; на поперечном срезе она выглядит как кольцо, названное образовательным, из которого закладывается стеблевой прокамбий. Протодерма и основная меристема классифицируются как полумеристемы: их клетки крупнее, более вакуолизированы и способны к растяжению, однако сохраняют делительную способность, обеспечивая утолщение формирующегося стебля.
Первичное утолщение стебля может принимать две формы. При кортикальном утолщении деление происходит в зоне первичной коры (как у подорожника), тогда как медуллярное утолщение связано с активным делением клеток будущей сердцевины (характерно для недотроги). Оба вида отличаются от вторичного утолщения, возникающего под действием камбия.
У семенных растений прокамбий возникает в основании листового зачатка: образуется очаг делящихся клеток, которые вытягиваются в длину, становятся прозенхимными и заострёнными на концах. Его развитие идет в двух направлениях: акропетально к верхушке листового бугорка и базипетально вниз по коровой части стебля, при этом прокамбий пересекает стебель под углом к продольной оси.
Камбий и феллоген относятся к боковым (латеральным) меристемам и участвуют в утолщении осевых органов. Камбий образуется из клеток прокамбия при их делении периклинальными перегородками. В отличие от прокамбия, формирующего первичные проводящие ткани, производные камбия дифференцируются во вторичные: наружные клетки образуют вторичную флоэму, внутренние — вторичную ксилему. Добавочный камбий может возникать не только из прокамбия, но и из дедифференцированных постоянных тканей, сохраняя структуру и функции обычного камбия. Феллоген (пробковый камбий) также образуется из постоянных тканей; при периклинальном делении наружные клетки становятся феллемами (пробковыми), а внутренние — клетками феллодермы.
У злаков (например, у ржи) инициальные клетки расположены на верхушке листового примордия, тогда как у большинства растений они находятся субапикально. Межпучковый камбий проявляет разную активность: у зонтичных он работает слабо, иногда образуя мелкие неполные пучки лишь из вторичной флоэмы; у лютиковых он не закладывается, а деятельность пучкового камбия почти отсутствует. У кирказона межпучковый камбий формирует паренхиму широких сердцевинных лучей, а у тыквенных в зрелых стеблях он отсутствует, и проводящие пучки имеют биколлатеральный характер.
Эволюция проводящей системы и стелярная теория
По мнению В. Циммермана, ключевым этапом эволюции наземных растений стало усложнение первичной ксилемы и флоэмы, приведшее к их дифференциации на прото- и метаксилему, а также на прото- и метафлоэму. Это позволило отказаться от простейших сосудистых систем и перейти к специализированным типам. Изначально заложение ксилемы было центрархным (сосуды в центре). Позже возник мезархный тип (сосуды в средней части), затем экзархный (ближе к периферии) и, наконец, эндархный, при котором расположение сосудов меняется в зависимости от доминирующих функций стебля.
Появление эндодермы у примитивных наземных растений стало важным барьерным адаптивным шагом: стенки её клеток, усиленные перицелем и утолщениями, препятствовали потере влаги. Однако с развитием устьичного аппарата и проводящей системы у семенных растений барьерная функция эндодермы утратилась, и в надземных стеблях она сохранилась лишь как крахмалоносное влагалище.
Параллельно шла дифференциация проводящей системы на осевую и листовую. Эволюционный тренд был направлен на редукцию осевой ткани и усиление листовых путей, что ярко проявилось у покрытосеменных: у однодольных почти вся проводящая система представлена листовыми следами. У микрофилльных растений (плауновидных) ключевым преобразованием стало дробление поперечных сечений ксилемы на тяжи. Современные плауноиды утратили типичную гаплостелу; её сменяют актиностела и плектостела — модификации протостелы. У редкого плауна ксилема имеет лопастное или почти звёздчатое строение, окруженное флоэмой (актиностела), а у булавовидного плауна ксилема расчленена на тяжи с экзархной протоксилемой, где флоэма заполняет промежутки, образуя плектовую структуру.
У папоротников (макрофиллов) наблюдается сифоностела — трубчатая структура. В корневищах сохраняется примитивная гаплостела, а в узлах листовых следов образуются заполненные паренхимой лакуны, придающие поперечному срезу подковообразную форму. В междоузлиях ксилема образует сплошной цилиндр без лакун. Метаксилема развивается центростремительно, состоя из трахеид с лестничной поровостью и паренхимы. Ситовидные клетки флоэмы с мелкими ситовидными полями представляют самые широкопросветные элементы системы.
Эвстела, внешне схожая с сифоностелой, отличается способностью к вторичному утолщению за счёт камбия и наличием сердцевинных лучей. Внутри неё открытые проводящие пучки могут располагаться вокруг центрального цилиндра (пучковое строение) или образовывать сплошной цилиндр (непучковое). Коллатеральные пучки характеризуются боковым расположением ксилемы рядом с флоэмой. У двудольных (например, тыквенных) встречаются биколлатеральные пучки, где ксилема лежит между двумя тяжами флоэмы. В корнях используется радиальное расположение, а в концентрических пучках одна ткань окружает другую: амфивазальные (ксилема вокруг флоэмы, у некоторых однодольных) и амфикрибральные (флоэма вокруг ксилемы, у папоротников).
Эволюция стел шла от простейшей гаплостелы с центрархной протоксилемой к сложным типам, оптимизируя соотношение объёма и площади проводящих тканей. Стелярная теория, основанная на трудах Ф. ван Тигема, А. Дулио, Ф. Боуэра, Ч. Джеффри, Г. Бребнера и В. Циммермана, подчёркивает существенные различия в структуре проводящей системы между отделами высших растений, связывая их с эволюционным прогрессом.
Анатомия стебля однодольных растений
У древоподобных однодольных (драцена, юкка, цветоносы агавы) наблюдается постоянное утолщение стеблей. Эта особенность поддерживается меристематической зоной, онтогенетически связанной с меристемой первичного утолщения. Зона образуется под конусом нарастания и у зрелых особей располагается между первичной корой и атактостелой, где пучки листовых следов распределены диффузно. Меристематический фронт формирует соединительную паренхиму, а пучки приобретают концентрическую амфивазальную организацию, отличающуюся от коллатеральных пучков большинства сосудистых растений.
Организация проводящих тканей у таких однодольных весьма разнообразна. Внутренние листовые пучки могут быть как коллатеральными, так и концентрическими, что определяется первичной стелярной организацией и вторичными изменениями, связанными с работой латеральных меристем. Эти анатомические варианты напрямую влияют на механические свойства стебля и его способность к росту.
В целом у однодольных пучки распределены по поперечному сечению хаотично, без строгой организации. У спаржи ксилема имеет V-образные очертания, а у рогоза ксилема и флоэма собраны в группы, вытянутые в тангенциальном направлении (метаксилема рогоза состоит лишь из двух широкопросветных сосудов). У некоторых пальм метаксилема представлена одним сосудом или широкой трахеидой, над флоэмой развиваются группы протофлоэмных волокон, а у сусаков и традесканций протоксилема разрушается на ранних этапах, образуя водоносный канал. Часто встречаются концентрические амфивазальные пучки (как в корневище ландыша); межпучковая паренхима может включать идиобласты, а у ароидных (аронник, колоказий) — млечники. У водных растений стелярная паренхима дифференцируется в аэренхиму. Перицикл хорошо развит у купены, лука, тюльпана и спаржи, представляя собой многослойную одревесневшую склеренхиму, иногда содержащую мелкие проводящие пучки.
Адаптации и специфические формы стеблей
Анатомическое разнообразие стеблей отражает широкий спектр экологических адаптаций. У лидирующей группы лиан атипичная стеблевая архитектура обеспечивает гибкость, необходимую для обвивания опор. Она основана на развитой паренхиме и ограниченном развитии механических тканей, что позволяет изгибаться без разрушения. У тропических лиан камбий часто работает неравномерно: в некоторых секторах образуется только луб, который затем погружается в древесину (как у энтады). У бигнонии это приводит к сокращению или прекращению образования древесины на отдельных участках с одновременным усилением образования луба, из-за чего срез приобретает лопастную форму.
У водных растений форма стебля варьируется от округлой до сплюснутой, от 3–4-гранной до бороздчатой или крыльчатой. Крылья и бороздки образуются при разрастании оснований листьев, придавая побегу специфическую гидродинамическую форму.
В злаковых и осоковых характерны воздухоносные полости, водоносные каналы, специфическая организация ксилемы и флоэмы с механическими обкладками, а также активное деление межпучковой паренхимы, способствующее восстановлению после перегиба. У злаков листовая система построена на принципе параллельного жилкования: все сосудисто-волокнистые пучки идут вдоль пластинки и сходятся к верхушке, где обычно располагается гидатода. Выделяют два типа листьев. Кранцевый лист отличается плотной паренхимной обкладкой из хлорофиллоносных клеток и радиально ориентированным мезофиллом. Обычный лист обладает механолимбной обкладкой (механическим влагалищем) из вытянутых одревесневших клеток. Склеренхима может образовывать тяжи по краям или сплошной тяж вдоль нижней стороны (как у ковыля), особенно развиваясь во влагалищах. У мятлика луковичного влагалища утолщаются за счёт запасающей паренхимы, а между пучками образуются крупные воздухоносные ходы, как у манника, облегчая газообмен и осмотический баланс.
Фототропизм иллюстрирует один из тропизмов — медленных движений, вызываемых неоднородным воздействием света. Электрофизиологическая поляризация приводит к асимметричному мембранному транспорту ауксина, концентрирующегося на освещенной стороне. На затенённой стороне клетки делятся быстрее, вызывая удлинение стебля и его выпрямление. В отличие от животных движений, механизм тропизмов основан не на мышечных сокращениях, а на гормональной и электрофизиологической регуляции.
Палеозойские голосеменные (семенные папоротники) с высокими стволами и крупными вайями демонстрировали одновременную активность нескольких камбиев. В их стеблях медуллезы в основании были протостеличными с экзархной или мезархной протоксилемой; от стелы отходили ветвящиеся в листья мелкие протостелы, окруженные перициклом.
Заключение
Стебель растения представляет собой не статичную структуру, а динамичную систему, чья анатомия тесно связана с эволюционной историей и экологическими условиями. От примитивных протостел до сложных эвстел и адаптивных атипичных утолщений — проводящая система непрерывно оптимизировалась для эффективного транспорта воды и органических веществ. Разнообразие тканей — от защитной эпидермы до опорной колленхимы и фотосинтезирующей хлоренхимы — демонстрирует поразительную пластичность растений. Независимо от того, растёт ли дерево в тропиках, злак в степи или лиана в лесу, анатомия стебля всегда является безупречным решением природной инженерии, обеспечивающим выживание и развитие организма.