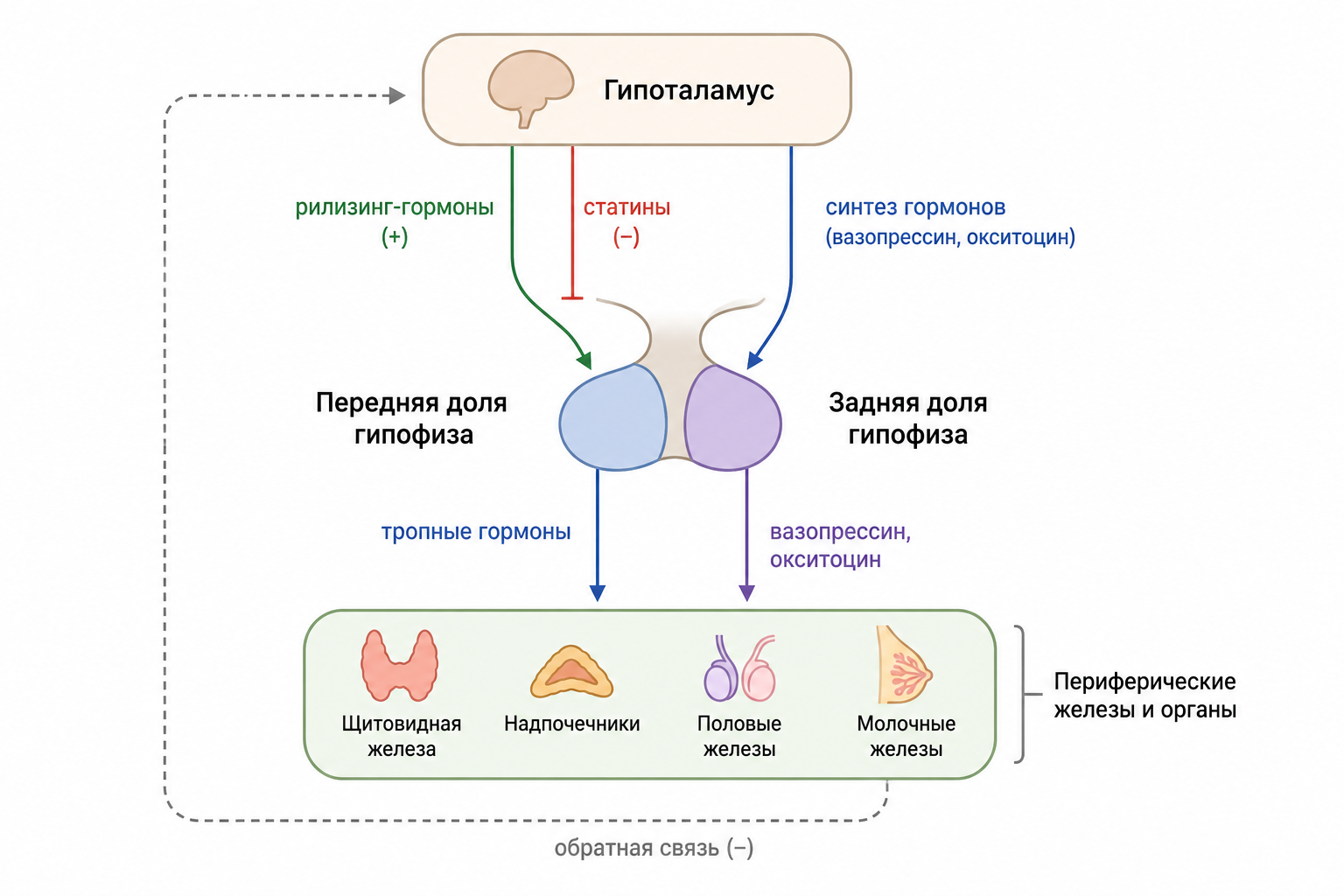
Гипофиз и его роль в эндокринной системе
Гипофиз — крошечная железа массой около полуграмма — управляет работой почти всей эндокринной системы организма. Несмотря на скромные размеры, он согласованно руководит ростом, обменом веществ, репродукцией и реакцией на стресс, превращая разрозненные сигналы гипоталамуса в точные гормональные команды. Понимание того, как устроены его доли, какие клетки вырабатывают гормоны и как эти сигналы достигают органов‑мишеней, позволяет увидеть единую архитектуру нейроэндокринной регуляции — от молекулярных механизмов синтеза до клинических синдромов при её нарушениях.
Передняя доля гипофиза: пять типов эндокринных клеток
Передняя доля гипофиза (аденогипофиз) образована гексагонально упакованными гормонсинтезирующими клетками, каждая из которых специализируется на выработке одного типа гормона. Выделяют пять морфологически различимых типов клеток — соматотропы, кортикотропы, тиреотропы, гонадотропы и лактотропы, — которые можно идентифицировать с помощью иммунохимических красителей, реагирующих с комплексом «антиген‑антитело», где антигеном выступает конкретный гормон.
Среди них различают ацидофильные клетки (соматотропы и лактотропы), базофильные клетки (кортикотропы и тиреотропы) и амфифильные гонадотропы, отличающиеся по электронно‑микроскопической структуре гранул. Соматотропы, отвечающие за синтез гормона роста (GH), составляют около 30–40 % общей численности клеток передней доли; они окрашиваются только кислыми красителями, за что и получили своё название, что в свою очередь обусловило термин «ацидофильные опухоли» для аденом, секретирующих избыточный GH. Кортикотропы, продуцирующие адренокортикотропный гормон (АКТГ), занимают приблизительно 20 % клеточного состава передней доли. Остальные три типа — тиреотропы, гонадотропы и лактотропы — вместе представлены менее чем 3–5 % каждый, однако секретируют чрезвычайно активные гормоны, контролирующие функции щитовидной железы, половые функции и лактацию.
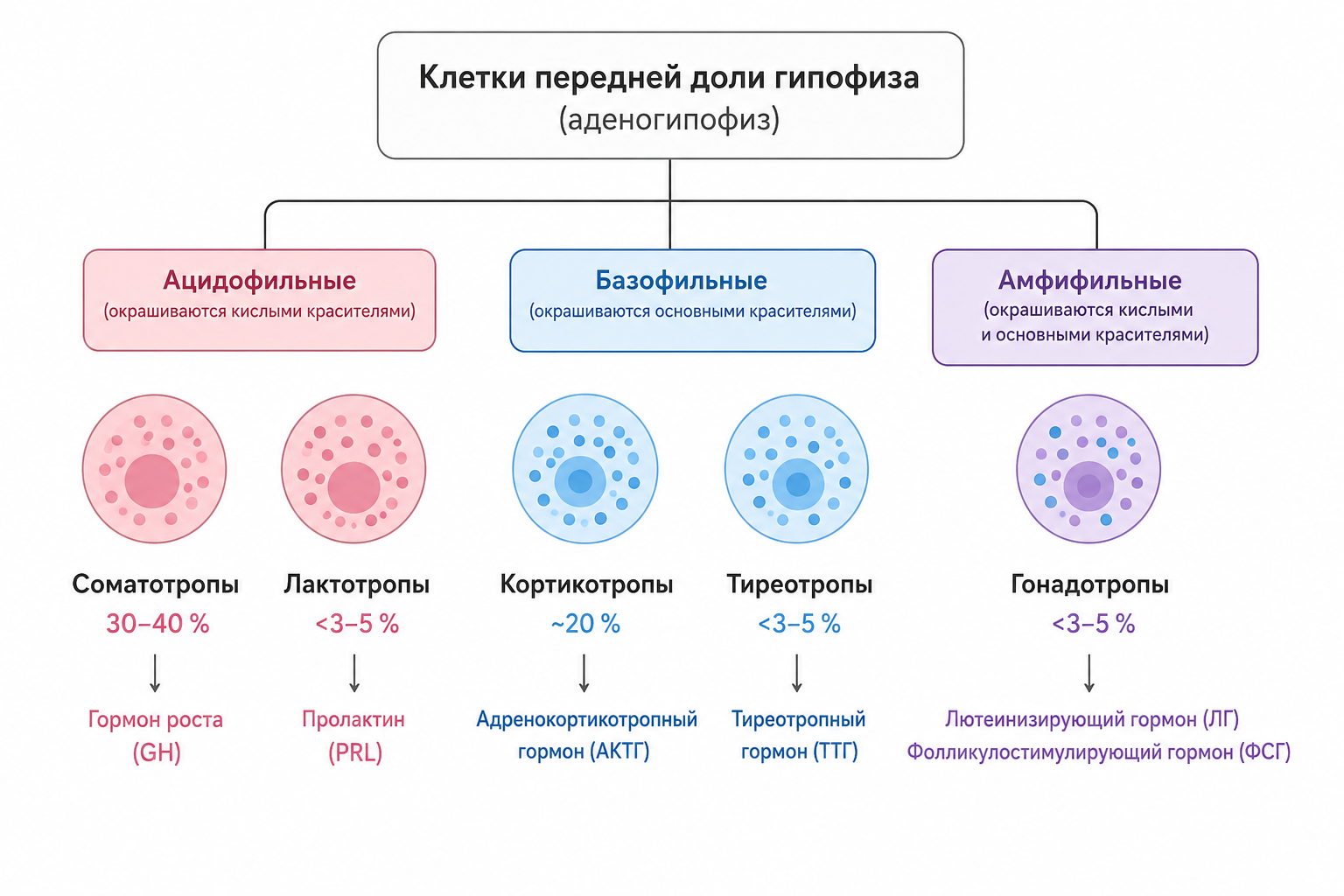 Рис. 1. Типы клеток передней доли гипофиза
Рис. 1. Типы клеток передней доли гипофиза
Синтез, упаковка и хранение гормонов
Клетки передней доли синтезируют гормоны в аппарате Гольджи, где предшественники гормонов обрабатываются ферментами, превращаясь в биологически активные пептиды; готовые гормоны упаковываются в секреторные везикулы, которые удерживаются мембраной в цитоплазме до стимуляции высвобождения. Все шесть основных гормонов передней доли — гормон роста, АКТГ, ТТГ, ФСГ, ЛГ и пролактин — являются пептидными (или белковыми), что определяет механизм их действия через мембранные рецепторы клеток‑мишеней. В венах передней доли эти гормоны собираются и поступают в общий кровоток для воздействия на соответствующие органы.
Задняя доля гипофиза и её гормоны
Гормоны задней доли — антидиуретический гормон (вазопрессин) и окситоцин — не синтезируются в самом гипофизе. Они образуются нейронами гипоталамуса в супраоптическом и паравентрикулярном ядрах, после чего транспортируются по аксону через гипофизарный нож (гипофизарный ствол) в заднюю долю, где хранятся до высвобождения в кровь. Структурно задняя доля (нейрогипофиз) состоит из крупных нейрональных окончаний, образующих сеть сосудистых синусов, что позволяет гормонам быстро попадать в системный кровоток — в отличие от более сложной капиллярной сети передней доли, организованной гипоталамо‑гипофизарной портальной системой.
В эмбриональном развитии передняя доля гипофиза происходит из гипофизарного протуберанса (рожка) эпителия, тогда как задняя доля формируется из выроста нейрональной ткани гипоталамуса. Это отражает их различный морфогенетический и клеточный источник: эпителиальный у аденогипофиза и нервный у нейрогипофиза.
Гипоталамо‑гипофизарная портальная система
Анатомически гипофиз соединён с гипоталамусом через короткую, но прочную артериально‑венозную систему. Кровь, поступающая в гипофизарный ствол, проходит сначала через капилляры гипоталамуса, затем через промежуточные сосуды гипофизарной портальной системы и наконец попадает в сеть синусов задней доли, обеспечивая две отдельные, но взаимосвязанные гормональные регуляторные схемы. Передняя доля получает регуляторные гипоталамические гормоны — тиреотропин‑рилизинг гормон, кортикотропин‑рилизинг гормон, гормон роста‑рилизинг гормон, гонадотропин‑рилизинг гормон и дофамин — через плотную сеть портальных сосудов; эти гормоны доставляются из гипоталамических ядер к соответствующим клеткам передней доли без прямого нервного контакта.
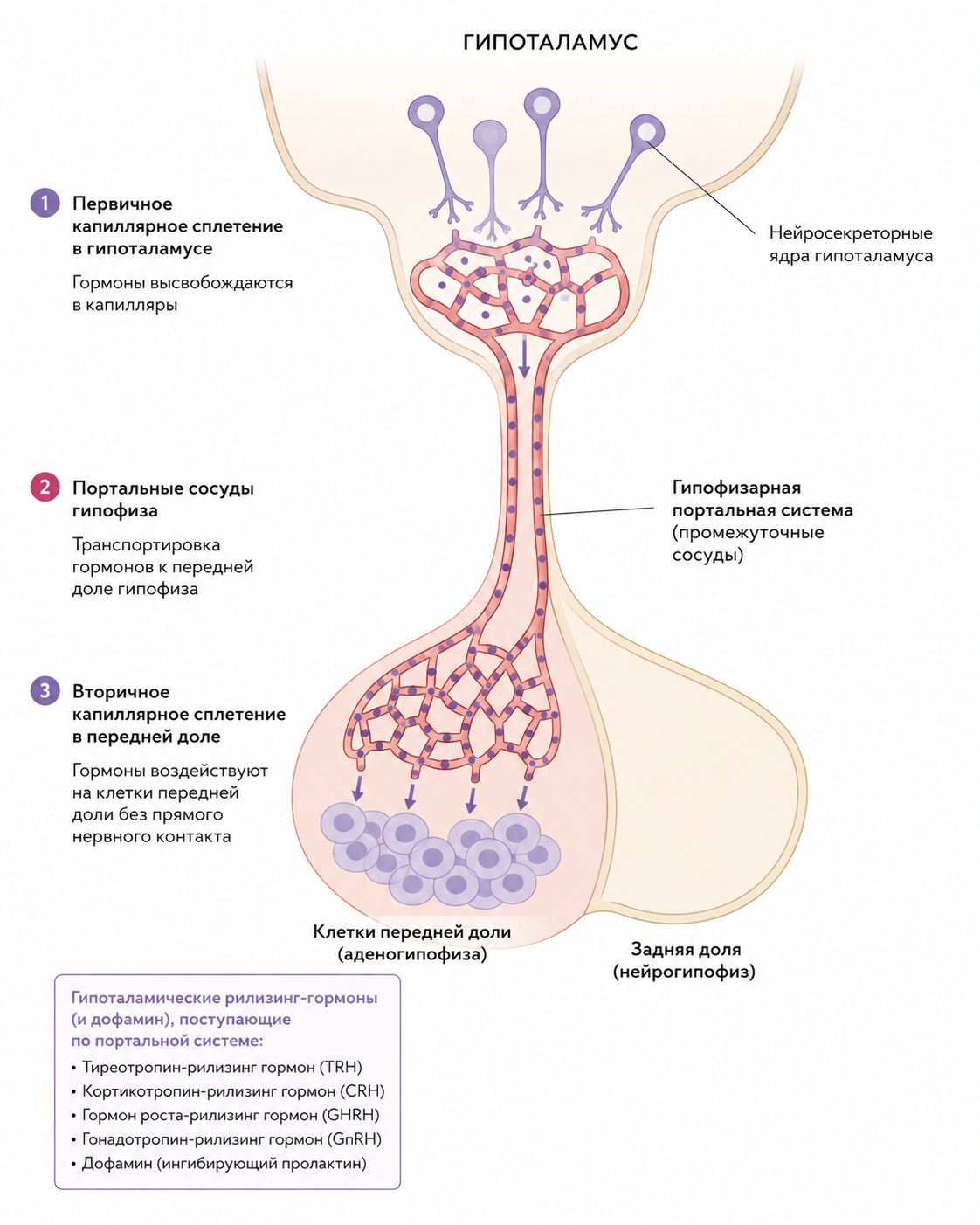 Рис. 2. Схема гипоталамо‑гипофизарной портальной системы, показывающая путь гипоталамических гормонов к клеткам передней доли гипофиза
Рис. 2. Схема гипоталамо‑гипофизарной портальной системы, показывающая путь гипоталамических гормонов к клеткам передней доли гипофиза
Гипоталамус как высший эндокринный регулятор
Гипоталамус составляет менее 1 % мозговой массы, однако является одним из важнейших регулирующих образований лимбической системы и контролирует большинство вегетативных и эндокринных функций организма. Он имеет двусторонние связи со всеми уровнями лимбической системы и посылает сигналы к стволу мозга, переднему таламусу, лимбическим частям коры, а также к воронке гипоталамуса для регуляции секреторных функций заднего и переднего гипофиза.
Секреция гормонов передней доли в значительной степени опосредована гипоталамическими рилизинг‑ и ингибирующими гормонами, которые выделяются нервными окончаниями срединного возвышения гипоталамуса и транспортируются в аденогипофиз. Электростимуляция этой области вызывает возбуждение нервных окончаний с последующим выделением практически всех гипоталамических гормонов, что подтверждает нейрогенную природу их секреции. Тела нервных клеток, дающих начало волокнам срединного возвышения, локализованы в других отделах гипоталамуса или ближайших базальных областях мозга, а не в самом срединном возвышении; их аксоны формируют проекции к срединному возвышению для высвобождения рилизинг‑гормонов.
Гипоталамические сигналы реализуются через две магистральные линии связи. Первая — продольный пучок, идущий от коры большого мозга вниз через середину гипоталамуса к ретикулярной формации ствола мозга и несущий волокна в обоих направлениях. Вторая — короткие пути между ретикулярной формацией мозгового ствола, таламусом, гипоталамусом и большинством других прилегающих областей основания мозга.
Гипоталамические рилизинг‑ и ингибирующие гормоны
Гормоны передней доли реализуют своё влияние преимущественно путём стимуляции желез‑мишеней — щитовидной железы, коры надпочечников, семенников, яичников и молочных желёз. Исключением является гормон роста, который, в отличие от других гормонов аденогипофиза, действует непосредственно на все ткани организма, а не через промежуточные железы‑мишени. Гипоталамус регулирует секрецию передней доли через рилизинг‑гормоны и ингибирующие гормоны, транспортируемые по портальной системе кровеносных сосудов.
Тиреотропин‑рилизинг гормон (ТРГ) — трипептид из трёх аминокислот — вызывает высвобождение тиреотропного гормона; его секреция подвержена влиянию низких температур и эмоциональных реакций. Кортикотропин‑рилизинг гормон (КРГ) — цепочка из 41 аминокислоты — стимулирует продукцию АКТГ; помимо эндокринной функции, он относится к анорексигенным факторам, снижающим потребление пищи. Гормон роста‑рилизинг гормон (ГРРГ) — цепочка из 44 аминокислот — усиливает выброс гормона роста. Гонадотропин‑рилизинг гормон (ГнРГ) — пептид из 10 аминокислот — стимулирует выделение двух гонадотропных гормонов, ЛГ и ФСГ; таким образом, один гипоталамический гормон контролирует продукцию двух аденогипофизарных гормонов, регулирующих функцию половых желёз.
Торможение секреции гормона роста осуществляется гормоном роста‑ингибирующим гормоном (соматостатином) — цепочкой из 14 аминокислот, обеспечивающим двустороннюю регуляцию. Торможение секреции пролактина выполняет пролактин‑ингибирующий гормон, которым является дофамин (катехоламин) — единственный гипоталамический ингибитор из числа катехоламинов, а не пептидов. Существуют также другие гипоталамические факторы, включая стимуляторы секреции пролактина, что указывает на сложную многофакторную регуляцию продукции этого гормона.
Механизмы отрицательной обратной связи
Увеличение содержания гормонов щитовидной железы в крови снижает продукцию тиреотропного гормона передней долей гипофиза. Если уровень тиреоидных гормонов превышает норму в 1,75 раза, продукция ТТГ падает практически до нуля, что демонстрирует мощность механизма отрицательной обратной связи. Почти все регуляторные проявления этого механизма сохраняются даже при отделении аденогипофиза от гипоталамуса, что указывает на прямое ингибирующее влияние тиреоидных гормонов на переднюю долю гипофиза. Благодаря этому концентрация свободных тиреоидных гормонов в крови поддерживается почти на постоянном уровне.
Низкие температуры и эмоциональные реакции увеличивают выброс ТТГ и тиреотропин‑рилизинг гормона, повышая продукцию тиреоидных гормонов. При этом ни эмоциональные факторы, ни низкие температуры не эффективны на фоне перерезки гипофизарного стебля, что подтверждает опосредованность этих реакций именно через гипоталамус.
Гормон роста (соматотропин)
Соматотропный гормон (гормон роста, соматотропин) представляет собой белковую молекулу, содержащую 191 аминокислотный остаток в единственной полипептидной цепи, с молекулярной массой 22 005 дальтон. Он стимулирует рост практически всех тканей организма, увеличивая как размеры клеток, так и количество митотических делений с последующей дифференцировкой — особенно в растущей кости и мышечной ткани. Эксперимент на крысах одного помёта показал, что ежедневное введение гормона роста приводит к более значительным привесам у подопытных животных по сравнению с контрольной группой, причём разница в массе тела становится заметной уже в первые дни жизни и сохраняется после окончания периода введения гормона.
Гормон роста осуществляет своё действие двумя путями: непосредственно воздействуя на все ткани организма и опосредованно — через стимуляцию секреции целевых желёз (щитовидной железы, коры надпочечников, семенников, молочных желёз) их собственных гормонов. Его секреция регулируется гипоталамусом: соматотропин‑рилизинг гормон (пептид из 44 аминокислотных остатков) усиливает выброс соматотропинами передней доли, тогда как соматостатин (пептид из 14 аминокислот) подавляет эту секрецию, обеспечивая двустороннюю гормональную регуляцию. Секреция гормона роста носит волнообразный характер и осуществляется преимущественно в фазу глубокого сна продолжительностью до 12–14 часов в сутки, что отражает специфический гипоталамический тип регуляции.
Адренокортикотропный гормон
Адренокортикотропный гормон (АКТГ, кортикотропин) образуется из проопиомеланокортина (ПОМК) путём процессинга прогормонконвертазой 1 (ПК1), которая представлена именно в аденогипофизе. На начальном этапе АКТГ стимулирует активацию протеинкиназы А, что приводит к превращению холестерола в прегненолон — лимитирующему по скорости этапу в синтезе всех адренокортикальных стероидных гормонов. Длительная стимуляция коры надпочечников АКТГ не только активирует секреторные процессы, но и приводит к гипертрофии и пролиферации клеток коры надпочечников, главным образом пучковой и сетчатой зон — именно эти зоны продуцируют кортизол и андрогены.
Физиологический стресс (болевые стимулы, физическое или умственное напряжение) в течение нескольких минут способен вызвать чрезвычайно сильное увеличение секреции АКТГ — иногда до 20 раз по сравнению с нормальным уровнем, соответственно возрастает и секреция кортизола. Болевые стимулы передают сигналы через ствол мозга в область срединного возвышения гипоталамуса, где образуется кортикотропин‑рилизинг‑гормон, поступающий в гипоталамо‑гипофизарную портальную систему. АКТГ специфически действует на кору надпочечников, стимулируя продукцию глюкокортикоидов — это пример прямой кросс‑регуляции, при которой гормон гипофиза целенаправленно контролирует секрецию гормонов периферической железы.
Тиреотропный, гонадотропные гормоны и пролактин
Тиреотропный гормон (тиреотропин, ТТГ), секретируемый тиреотропами, контролирует скорость секреции тироксина (Т4) и трийодтиронина (Т3) щитовидной железой; эти тиреоидные гормоны регулируют скорость основных химических реакций, поэтому увеличение продукции тироксина повсеместно усиливает обмен глюкозы и приводит к повышенной продукции инсулина поджелудочной железой.
Гонадотропины вырабатывают два гонадотропных гормона — фолликулостимулирующий гормон (ФСГ) и лютеинизирующий гормон (ЛГ). ФСГ стимулирует рост фолликулов в яичнике и сперматогенез в семенниках, а ЛГ вызывает овуляцию и стимулирует продукцию половых гормонов. Регуляция их секреции осуществляется гипоталамусом через гонадотропин‑рилизинг гормон.
Пролактин, продуцируемый лактотропами (маммотропами), обеспечивает развитие молочных желёз и продукцию молока, стимулируя лактацию после родов. Его секреция тормозится дофамином — пролактин‑ингибирующим гормоном, который подавляет выработку пролактина в отсутствие стимуляции сосания. Напротив, тиреотропин‑рилизинг гормон стимулирует секрецию не только ТТГ, но и пролактина, что объясняет повышение пролактина при первичном гипотиреозе, когда уровень ТРГ растёт в ответ на дефицит тиреоидных гормонов.
Взаимодействие эндокринных осей
Тиреоидные гормоны играют центральную роль в согласовании работы сразу нескольких периферических желёз. В механизме отрицательной обратной связи они повышают скорость инактивации глюкокортикоидов в печени, что, согласно принципу обратной связи, стимулирует повышенную продукцию АКТГ передней долей гипофиза и последующее увеличение выработки глюкокортикоидов корой надпочечников — это пример косвенного влияния щитовидной железы на функцию надпочечников через гипофизарно‑надпочечниковую ось.
Помимо этого, тиреоидные гормоны интенсифицируют обмен глюкозы, вызывая компенсаторное повышение секреции инсулина поджелудочной железой, что иллюстрирует их способность стимулировать активность большинства других эндокринных желёз и одновременно увеличивать тканевую потребность в гормонах. Они также усиливают метаболические процессы, связанные с формированием костей, что повышает потребность в паратиреоидном гормоне (ПТГ) паратиреоидных желёз — показательный пример того, как активация одной эндокринной оси (щитовидной железы) через изменение метаболических потребностей косвенно стимулирует другую (паратиреоидную).
Тиреоидные гормоны и половая функция
Для сохранения нормальных половых функций уровень тиреоидных гормонов должен оставаться в пределах нормы. У мужчин дефицит этих гормонов, по‑видимому, приводит к потере либидо, а их избыток может вызывать импотенцию — это указывает на тесную связь между функцией щитовидной железы и половыми железами. У женщин гипотиреоз часто проявляется меноррагией или полименореей — чрезмерно обильными и учащёнными менструациями, иногда сопровождаясь нарушением цикла вплоть до аменореи, а также значительным снижением либидо. При гипертиреозе у женщин наблюдается олигоменорея — очень скудные менструации — и иногда аменорея. Непосредственного влияния тиреоидных гормонов на гонады не выявлено; вероятна комбинация прямого метаболического воздействия на половые железы и опосредованного регулирования через гипофиз.
Гормоны яичников, надпочечников и интеграция с нервной системой
Гормоны яичника оказывают специфическое действие на женские половые органы и развитие вторичных половых признаков, реализуя эффекты, инициированные гонадотропными гормонами гипофиза, и замыкая тем самым петлю обратной связи. В то же время кора надпочечников вырабатывает небольшие количества андрогенов, которые при патологической гиперпродукции могут вызывать маскулинизацию организма, что демонстрирует функциональное перекрытие между надпочечниками и половыми железами в секреции андрогенов.
Мозговое вещество надпочечников и гипофиз реагируют на нервные стимулы, иллюстрируя интеграцию нервной и эндокринной систем. Мозговое вещество функционально относится к симпатическому отделу автономной нервной системы и секретирует адреналин и норадреналин в ответ на симпатическую стимуляцию, вызывая эффекты, схожие с прямой симпатической активацией. Аксоны нейроэндокринных клеток гипоталамуса оканчиваются в задней доле гипофиза и срединном возвышении, выделяя антидиуретический гормон, окситоцин и тропные гормоны, регулирующие секрецию передней доли. Тропные гормоны (тиреотропин‑рилизинг гормон и другие) служат связующим звеном между гипоталамусом и гипофизом, обеспечивая многоуровневую регуляцию периферических желёз.
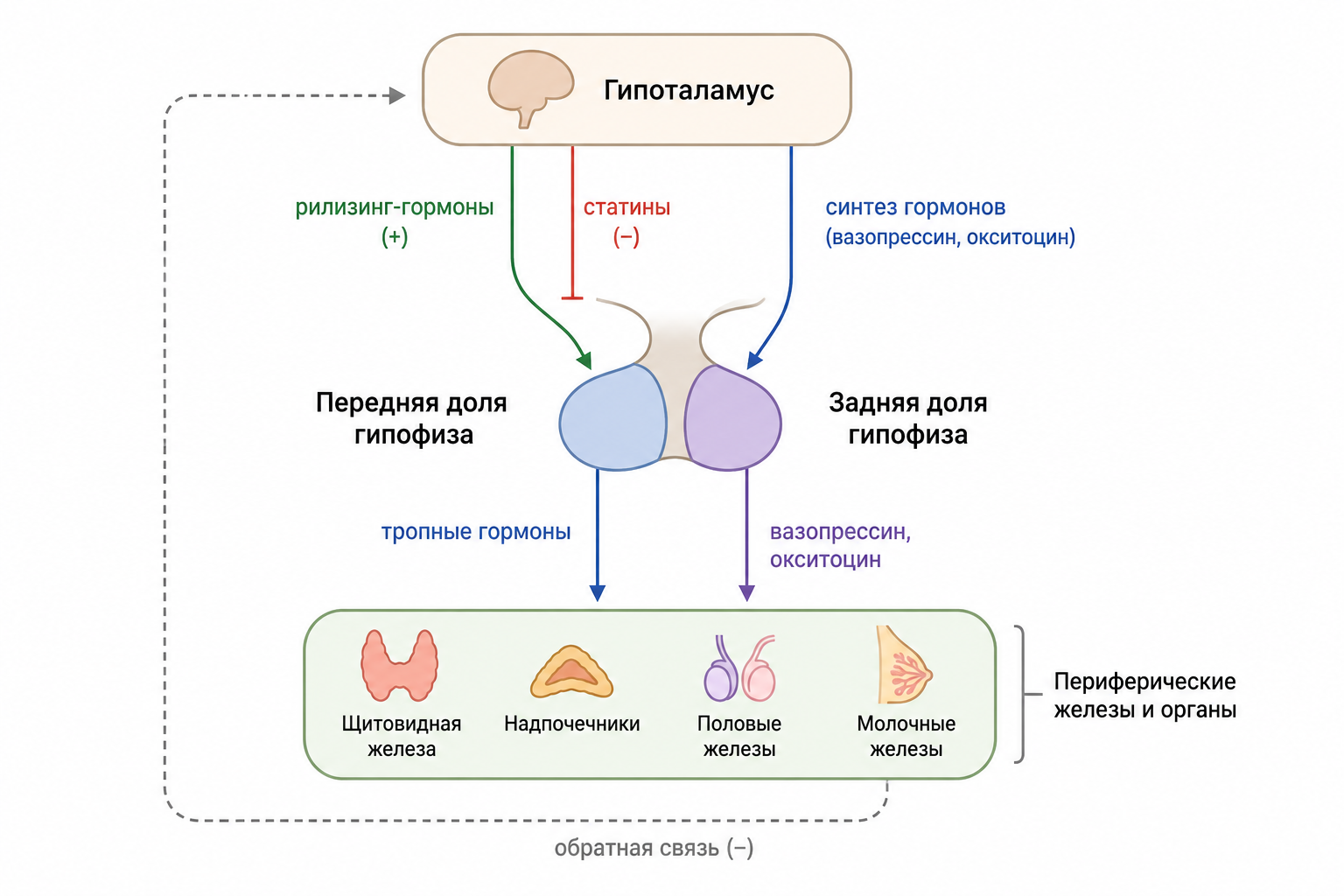 Рис. 3. Гипоталамо-гипофизарная ось (Схема взаимодействия гипоталамуса, передней и задней доли гипофиза и периферических желез).
Рис. 3. Гипоталамо-гипофизарная ось (Схема взаимодействия гипоталамуса, передней и задней доли гипофиза и периферических желез).
Доставка гормонов к тканям и тканевая специфичность
Эндокринные гормоны транспортируются кровью ко всем клеткам организма, включая нервную систему, где они связываются с рецепторами и запускают разнообразные реакции. Некоторые гормоны — такие как гормон роста и тироксин — действуют на широкий спектр клеток, стимулируя рост или ускоряя химические реакции. Другие воздействуют только на специфические ткани‑мишени, поскольку соответствующие рецепторы присутствуют лишь в этих тканях. Плацента является дополнительным источником половых гормонов: она представляет собой временную эндокринную железу, обеспечивающую гормональную поддержку беременности и взаимодействующую с гипофизарно‑гонадной осью.
Гигантизм
Гигантизм развивается при резком повышении активности ацидофильных клеток аденогипофиза, продуцирующих гормон роста, что чаще всего обусловлено возникновением опухолевого процесса в этих клетках. Избыточная продукция соматотропина вызывает интенсивный рост всех тканей, включая костную; если это состояние манифестирует до закрытия эпифизарных зон роста трубчатых костей, рост индивидуума может достигать 2,5 м. Гигантизму сопутствует гипергликемия, развивающаяся вследствие гиперстимуляции бета‑клеток островков Лангерганса поджелудочной железы избыточной концентрацией глюкозы; приблизительно в 10 % случаев это приводит к развитию сахарного диабета. При отсутствии лечения гигантизм сменяется пангипопитуитаризмом, поскольку опухоль со временем вызывает распад аденогипофиза, что, как правило, приводит к ранней гибели; однако микрохирургическое лечение или облучение гипофиза способны предотвратить летальный исход.
Акромегалия
Акромегалия возникает при развитии ацидофильных опухолей гипофиза в зрелом возрасте, когда эпифизарные зоны роста трубчатых костей уже закрыты и рост в высоту невозможен. Вместо этого происходит пропорционально неравномерный рост мягких тканей и утолщение костей в ширину. Наиболее характерно разрастание губчатых костей черепа — нос, лобные бугры, надбровные дуги, нижняя челюсть, позвонки — а также рост кистей и стоп. При этом нижняя челюсть выдаётся вперёд более чем на 1 см, нос увеличивается приблизительно в 2 раза, а пальцы резко утолщаются. Вследствие разрастания тканей язык, печень и особенно почки значительно увеличиваются в размерах, а рост позвонков приводит к формированию кифоза.
Гипогонадизм у женщин
Гипогонадизм у женщин может быть следствием врождённого отсутствия яичников, генетически обусловленного дефекта ферментных систем, ответственных за синтез половых гормонов в клетках яичников, или недоразвития яичников. При врождённом отсутствии яичников или невозможности их функционирования с наступлением пубертатного периода формируется женский евнухоидизм, характеризующийся отсутствием вторичных половых признаков при сохранении детских характеристик половых органов, а также чрезмерно продолжительным ростом трубчатых костей из‑за позднего закрытия зон роста — в результате рост женщины с евнухоидизмом соответствует или даже превышает рост мужчины с аналогичными нарушениями. При задержке развития яичников наблюдается параллельная задержка развития вторичных половых признаков и внутренних половых органов: матка у взрослой женщины может соответствовать размерам детской, влагалище остаётся тонкостенным и легко повреждается, молочные железы атрофируются.
Нарушение циклической работы яичников вплоть до аменореи при гипогонадизме обусловлено тем, что для поддержания ритмичного полового цикла снижение продукции эстрогенов не должно опускаться ниже определённого критического уровня; при его достижении циклическая активность яичников нарушается. Аналогичные изменения половых органов — атрофия молочных желёз, матки, тонкость эпителия влагалища, скудное оволосение лобка — наблюдаются и в постменопаузальном периоде.
Болезнь Аддисона
Болезнь Аддисона (гипоадренализм) является результатом первичной атрофии коры надпочечников. В 80 % случаев она обусловлена аутоиммунным процессом, но может также возникать вследствие туберкулёзного поражения надпочечников или их деструкции опухолевого генеза. Дефицит минералокортикоидов приводит к резкому снижению канальцевой реабсорбции натрия в почках, что вызывает массивную потерю ионов натрия, хлоридов и воды с быстрым уменьшением объёма внеклеточной жидкости и развитием гипонатриемии, гиперкалиемии и лёгкого ацидоза. Снижение объёма плазмы крови ведёт к увеличению концентрации эритроцитов, уменьшению сердечного выброса и развитию шока; в нелеченных случаях смерть наступает в срок от 4 суток до 2 недель после прекращения продукции минералокортикоидов. Гипофункция коры надпочечников в значительной степени опосредована гипоталамическим ингибирующим гормоном, который снижает продукцию адренокортикальных и тиреоидных гормонов; состояние купируется введением адренокортикальных гормонов и гормонов щитовидной железы.
Внутриклеточные вторичные посредники
Гормон, связываясь с мембранным рецептором, активирует Gs‑белок, который в свою очередь стимулирует мембраносвязанную аденилатциклазу. Этот фермент превращает небольшое количество цитоплазматического АТФ в циклический АМФ (цАМФ), преобразуя гормональный сигнал во внутриклеточный вторичный посредник. Образование цАМФ запускает каскадную реакцию, в которой каждый активированный фермент активирует следующий, что усиливает ответ: ничтожно малое количество гормона способно активировать значительно большее количество молекул на последующих этапах. цАМФ реализует своё действие через активацию цАМФ‑зависимой протеинкиназы (ПКА), которая фосфорилирует специфические белки и запускает биохимические реакции клетки; непосредственное действие гормона ограничивается активацией рецептора, а все остальные процессы обеспечиваются вторичными посредниками.
Помимо цАМФ, важными внутриклеточными посредниками являются ионы кальция, связанные с кальмодулином, и фосфолипидные фрагменты мембран — диацилглицерол (ДАГ), инозитолтрифосфат (ИФ3) и фосфатидилинозитолбифосфат (ФИФ2), образующиеся при гидролизе мембранных фосфолипидов и участвующие в передаче сигнала внутрь клетки.
Синтез и секреция гормонов железистыми клетками
Секреторные белки‑гормоны синтезируются на рибосомах, прикреплённых к шероховатому эндоплазматическому ретикулуму, отвечающему за построение секретируемых белков. Далее они перемещаются по канальцам ЭР, преодолевая этот путь примерно за 20 минут, и достигают комплекса Гольджи. В аппарате Гольджи секреторные материалы модифицируются, сортируются, концентрируются и высвобождаются в цитоплазме в виде секреторных пузырьков, которые накапливаются на апикальном конце железистой клетки. Ферменты этих пузырьков расщепляют прогормоны, формируя биологически активные гормоны и неактивные фрагменты. Секреторные пузырьки хранятся в цитоплазме, многие из них остаются связанными с плазматической мембраной до тех пор, пока не поступит нервный или гуморальный сигнал, запускающий их экзоцитоз.
💯 Подробнее: экзоцитоз секреторных пузырьков в железистой клетке
Процесс экзоцитоза начинается с увеличения проницаемости клеточной мембраны для ионов кальция под воздействием управляющего сигнала; кальций поступает в клетку и способствует слиянию большого количества секреторных пузырьков с апикальной поверхностью мембраны. Затем апикальная мембрана разрывается, высвобождая содержимое пузырьков наружу — этот этап и называется собственно экзоцитозом.
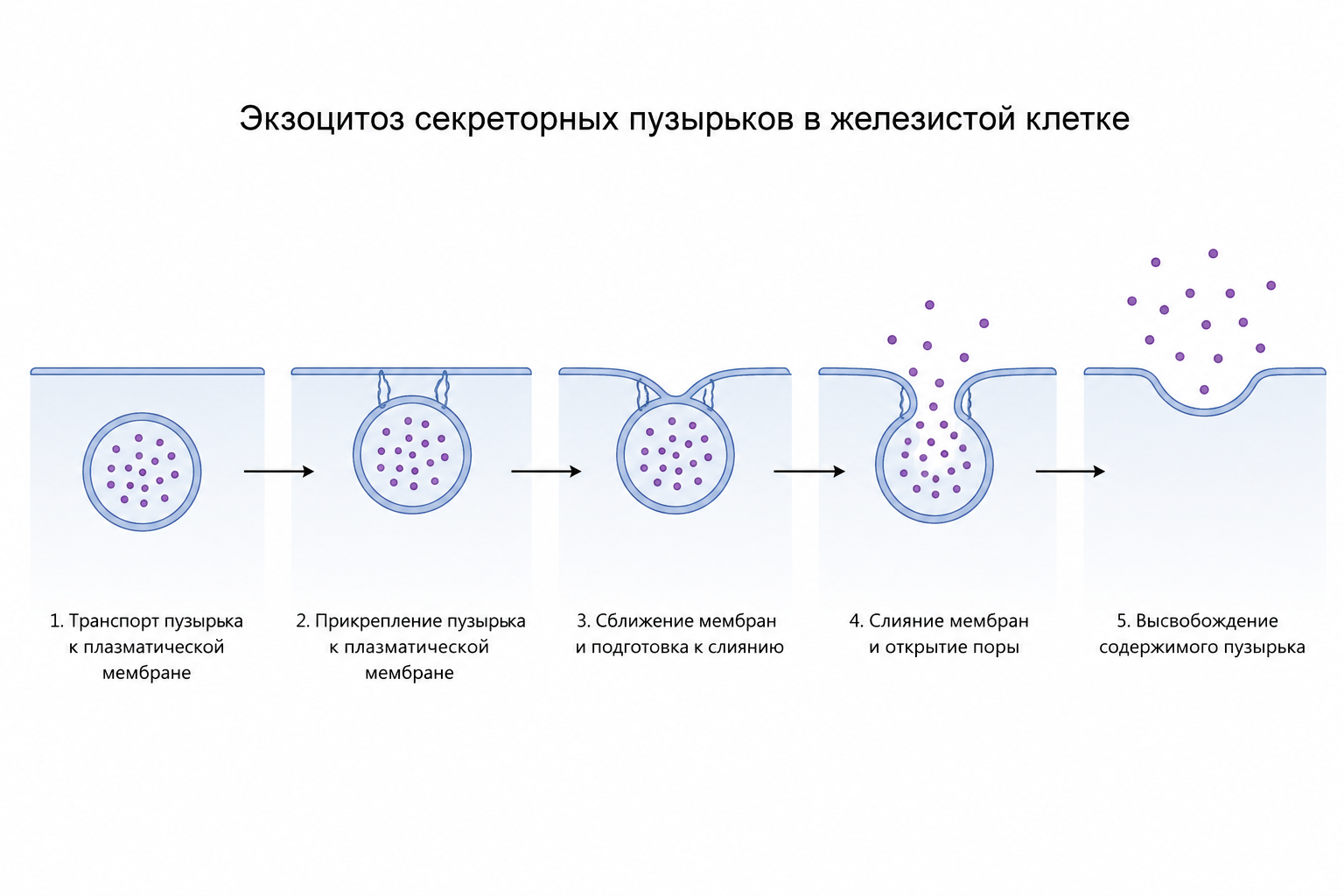 Рис. 4. Схема процесса экзоцитоза секреторных пузырьков в железистой клетке
Рис. 4. Схема процесса экзоцитоза секреторных пузырьков в железистой клетке
Сопряжённая секреция воды и электролитов
Помимо секреции гормонов, железистая клетка обеспечивает выход воды и электролитов, необходимых для сопровождения органических веществ. Нервная стимуляция оказывает специфическое действие на базальную клеточную мембрану, активируя активный транспорт ионов хлора внутрь клетки. Поступление хлора увеличивает электроотрицательность внутри клетки, что вынуждает положительные ионы — прежде всего натрий — двигаться через мембрану внутрь. Избыток отрицательных и положительных ионов создаёт осмотические силы, способствующие притоку воды, увеличению клеточного объёма и повышению гидростатического давления. Повышение давления приводит к открытию секреторной границы клетки, вызывая выход воды, электролитов и органических веществ из секреторного края железистой клетки. Подтверждением этой схемы служат расположение нервных окончаний у основания клеток и данные микроэлектродных исследований о наличии электрического мембранного потенциала на её базальной поверхности.
Итоговая карта гормонов гипофиза
Передняя доля гипофиза секретирует гормон роста, тиреотропный гормон, адренокортикотропный гормон, пролактин, фолликулостимулирующий гормон и лютеинизирующий гормон — все они являются пептидами. Задняя доля выделяет антидиуретический гормон (вазопрессин) и окситоцин. Секреция гормонов передней доли регулируется гипоталамическими рилизинг‑гормонами и ингибирующими гормонами, доставляемыми через гипоталамо‑гипофизарные портальные сосуды. К рилизинг‑гормонам относятся тиреотропин‑рилизинг гормон, кортикотропин‑рилизинг гормон, гормон роста‑рилизинг гормон и гонадотропин‑рилизинг гормон; к ингибирующим — гормон роста‑ингибирующий гормон (соматостатин) и дофамин (пролактин‑ингибирующий фактор).
Заключение
Гипоталамо‑гипофизарная система представляет собой многоуровневый регуляторный комплекс, в котором гипоталамус через рилизинг‑ и ингибирующие гормоны управляет передней долей гипофиза, а через нейрональные отростки — задней долей. Пять типов клеток аденогипофиза вырабатывают шесть пептидных гормонов, действующих либо непосредственно на ткани (гормон роста), либо через периферические железы‑мишени. Тесная взаимосвязь щитовидной оси, надпочечниковой оси и гонадной оси обеспечивается механизмами отрицательной обратной связи и общими внутриклеточными каскадами вторичных посредников — цАМФ, ионов кальция, ДАГ, ИФ3 и ФИФ2. Нарушения на любом уровне этой системы — от опухолей аденогипофиза (гигантизм, акромегалия) до аутоиммунной деструкции коры надпочечников (болезнь Аддисона) и гипогонадизма — демонстрируют, насколько точно должна быть сбалансирована секреция каждого гормона для поддержания гомеостаза.