Гипоталамус и его роль в эндокринной системе
Маленький, но невероятно мощный участок мозга — гипоталамус — является истинным дирижером нашей эндокринной системы. Располагаясь в самом центре лимбической структуры, он неразрывно связывает наши эмоции, поведенческие реакции и глубинные физиологические процессы, обеспечивая точную адаптацию организма к любым вызовам внутренней и внешней среды.
Анатомия и ядерная организация гипоталамуса
Гипоталамус занимает центральную позицию лимбической системы, находясь между передней спайкой, передними ядрами таламуса, гиппокампом, миндалиной и базальными ганглиями. Интеграция этой субчасти мозга с лимбической корой подтверждает его активную роль в связывании эмоциональных, поведенческих и физиологических сигналов.
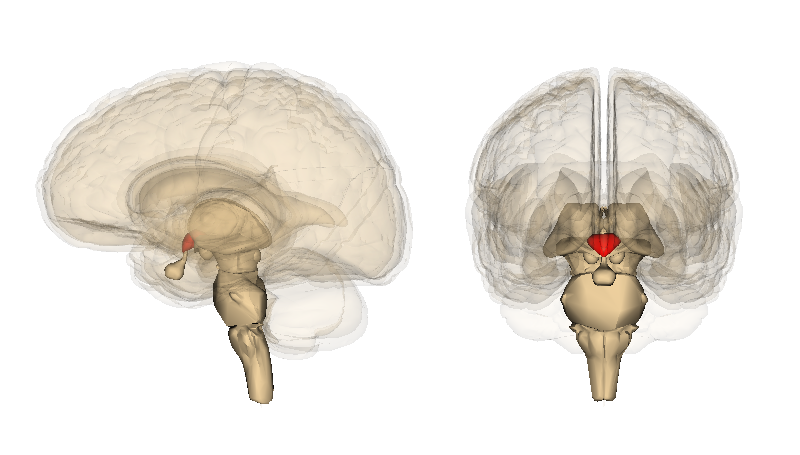 Рис. 1. Анатомическое расположение гипоталамуса. Гипоталамус обозначен красным цветом.
Рис. 1. Анатомическое расположение гипоталамуса. Гипоталамус обозначен красным цветом.
Images are generated by Life Science Databases(LSDB)., CC BY-SA 2.1 JP https://creativecommons.org/licenses/by-sa/2.1/jp/deed.en, via Wikimedia Commons
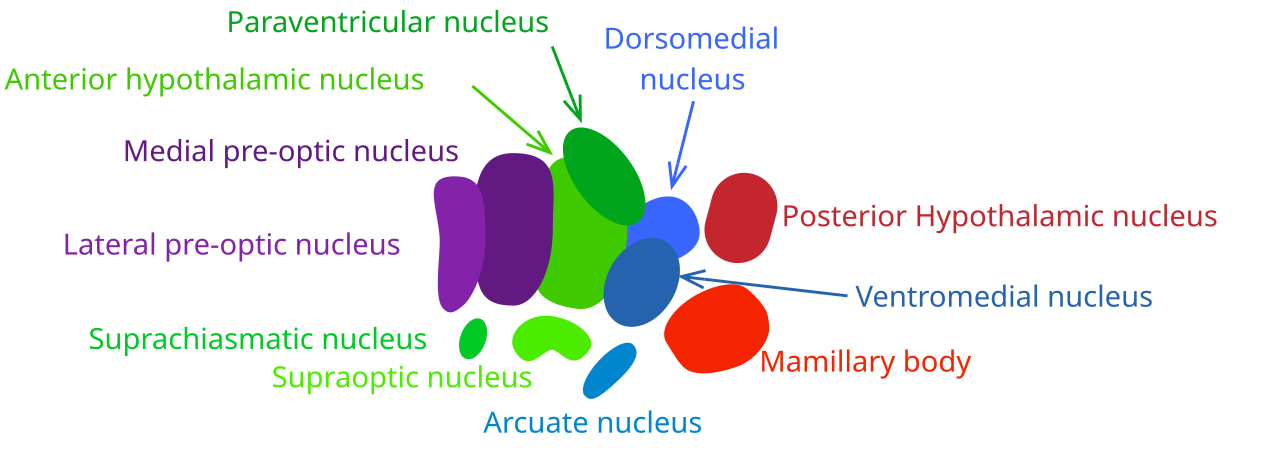
Внутри гипоталамуса выделяются несколько ядер, образующих коронарные и сагиттальные группы: передняя (преоптическая) зона, аркуатное ядро, медиальное (вентромедиальное) ядро, дорсомедиальное ядро, паравентрикулярное ядро, перивентрикулярная зона и дугообразное ядро. Каждый из этих центров имеет характерные морфологические границы и специфические нейрональные переходы.
Преоптическая (медиальная преоптическая) область, расположенная ближе к стенке третьего желудочка, контролирует сокращение мочевого пузыря, частоту сердцебиения и артериальное давление, участвуя в регуляции вегетативных функций нижних отделов мозга. Аркуатное ядро, находящееся вблизи дорсомедиального ядра, получает мультицитокиновый сигнал из желудочно-кишечного тракта и жировой ткани, координируя пищевое поведение, энергозатраты и гормональную регуляцию аппетита. Дорсомедиальное ядро отвечает за стимуляцию желудочно-кишечного тракта, повышение артериального давления и реакцию зрачков (расширение, дрожание), образуя часть традиционного «заднего гипоталамуса», связанного с симпатической активацией.
Вентромедиальное ядро, расположенное в передне-медиальной части гипоталамуса, считается центром насыщения: электрическая стимуляция этого ядра вызывает ощущение сытости и окончание пищевого поведения, а его разрушение приводит к непрекращающейся гиперфагии и массивному набору веса, иногда в четырёхкратном превышении нормы. Паравентрикулярное ядро, лежащее в непосредственной близости к третьему желудочку, регулирует секрецию окситоцина и удержание воды, участвуя в водно-солевом гомеостазе и социально-поведенческих реакциях, связанных с родами и лактацией. Сосцевидное тело гипоталамуса (соответствующее «пищевому» центру) находится в передне-дорсальной части и контролирует пищевые рефлексы, а также интегрирует соматосенсорные сигналы, связанные с жаждой и голодом. Перифорникальная зона участвует в генерации голодных сигналов, повышении артериального давления и яркости восприятия, предоставляя нейронные коррелаты мотивации к поиску пищи. Дугообразное ядро и прилегающая к нему перивентрикулярная зона формируют нейроэндокринный регуляторный центр, который синтезирует и высвобождает гипоталамические гормоны, влияющие на гипофизарную функцию и общую энергетическую гомеостазу.
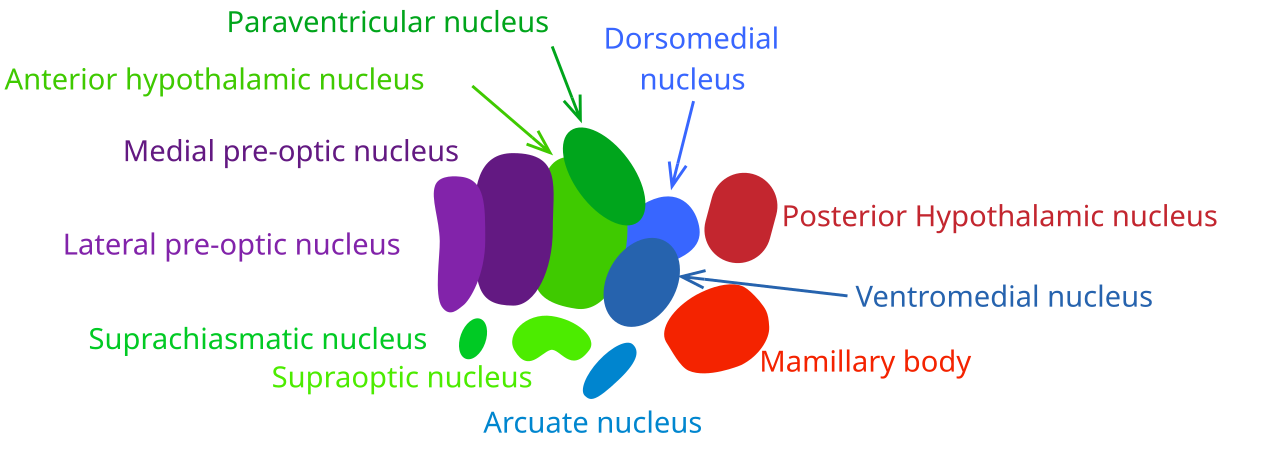 Рис. 2. Ядра гипоталамуса.
Рис. 2. Ядра гипоталамуса.
Oldblueday, CC BY-SA 4.0 https://creativecommons.org/licenses/by-sa/4.0, via Wikimedia Commons
Все упомянутые ядра соединены с лимбической корой, образующей кольцевое образование вокруг больших полушарий. Это кольцо включает орбитофронтальную, парагиппокампальную, зубчатую и поясную извилины, обеспечивая интеграцию сигналов. Гипоталамус образует центральный элемент в этом кольце, где каждая из извилин отсылает к нему через межконтурные проекции, создавая двойную связь между высшими когнитивными процессами и базовыми вегетативными регуляторами.
Рилизинг- и ингибирующие гормоны
Гипоталамические рилизинг- и ингибирующие гормоны образуют центральный регуляторный узел, контролирующий функции передней доли гипофиза. В их число входят тиреотропин-рилизинг гормон (Т-РГ), кортикотропин-рилизинг гормон (К-РГ), гормон роста-рилизинг гормон (Г-РГ), соматостатин (гормон роста-ингибирующий гормон), гонадотропин-рилизинг гормон (Гн-РГ) и дофамин, выступающий в роли пролактин-ингибирующего гормона. Каждый гормон передней доли находится под двойным контролем — стимуляторами и ингибиторами, что обеспечивает тонкую настройку эндокринных функций.
Структурное разнообразие этих факторов отражает их различное происхождение. Т-РГ представляет собой простейший трипептид (пироглутамил-гистидил-пролинамид), тогда как Гн-РГ состоит из десяти аминокислот, К-РГ — из 41, а Г-РГ — из 44. Соматостатин состоит из четырнадцати аминокислот. Дофамин, в отличие от всех перечисленных, не относится к пептидам, а является катехоламином, что делает его единственным непептидным гипоталамическим ингибирующим фактором. Разнообразие молекулярного строения определяет различные способы взаимодействия с рецепторами мишеней и биохимическую активность.
Гипоталамо-гипофизарная портальная система
Все гипоталамические гормоны синтезируются в нервных окончаниях срединного возвышения гипоталамуса и попадают в аденогипофиз по специальной гипоталамо-гипофизарной портальной системе. Тела нейронов, от которых отходят волокна срединного возвышения, расположены в иных гипоталамических ядрах и прилегающих базальных областях. Электростимуляция этой области приводит к высвобождению почти всех гипоталамических гормонов, что подтверждает её роль как центрального «переключателя» эндокринной регуляции. Портальная система обеспечивает доставку гормонов непосредственно к клеткам передней доли в высоких концентрациях, минуя общий кровоток; при её блокировке секреция тиреотропного гормона (ТТГ) резко снижается, хотя и сохраняется небольшая базальная активность, свидетельствующая о существовании дополнительных, менее значимых путей.
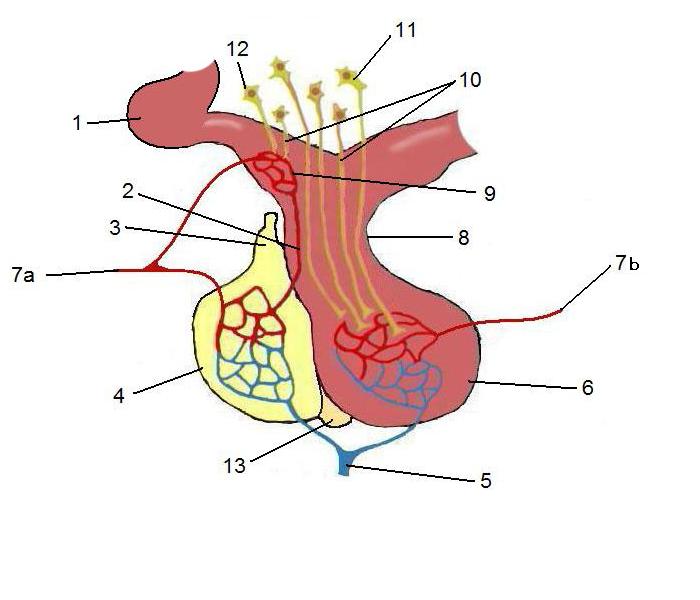 Рис. 3. Схема гипоталамо‑гипофизарной портальной (воротной) системы. 2 - гипофизарная воротная вена; 3 - гипофизарная вена; 4 - аденогипофиз (передний гипофиз); 6 - нейрогипофиз (задний гипофиз); 7a - верхняя гипофизарная артерия; 7b - нижняя гипофизарная артерия; 10 - нейросекреторные клетки: 11 - паравентрикулярного ядра и 12 - супраоптического ядра.
Рис. 3. Схема гипоталамо‑гипофизарной портальной (воротной) системы. 2 - гипофизарная воротная вена; 3 - гипофизарная вена; 4 - аденогипофиз (передний гипофиз); 6 - нейрогипофиз (задний гипофиз); 7a - верхняя гипофизарная артерия; 7b - нижняя гипофизарная артерия; 10 - нейросекреторные клетки: 11 - паравентрикулярного ядра и 12 - супраоптического ядра.
Takuma-sa, CC BY 3.0 https://creativecommons.org/licenses/by/3.0, via Wikimedia Commons
Молекулярный механизм действия Т-РГ на аденогипофиз иллюстрирует типичный путь трансдукции сигнала. Т-РГ связывается с высокоспецифичными рецепторами на мембране тиротрофов, что приводит к повышению внутриклеточного кальция и образованию диацилглицерола. Эти вторичные посредники активируют фосфолипазную систему, в частности фосфолипазу C, что в итоге усиливает синтез и высвобождение ТТГ. Подобные каскады, в которых вторичным посредником может быть цАМФ, характерны для многих тканей-мишеней, подчёркивая универсальность сигнального аппарата. Холодовая экспозиция служит ярким примером внешнего стимула, усиливающего продукцию Т-РГ: при воздействии низких температур активируются терморегуляторные центры, что повышает секрецию Т-РГ и увеличивает выброс ТТГ. Эксперименты с крысами, находившимися в экстремальном холоде в течение нескольких недель, демонстрируют многократный рост уровня тиреоидных гормонов.
Механизмы действия ключевых гормонов
Тиреотропин-рилизинг гормон (ТРГ) является главным гипоталамическим стимулятором секреции тиреотропного гормона (ТТГ) из тиреотропных клеток передней доли гипофиза. Его высвобождение происходит в ответ на снижение концентраций тиреоидных гормонов в плазме; образуется отрицательная обратная связь, при которой повышенный уровень ТТГ подавляет дальнейшую выработку ТРГ.
Гонадотропин-рилизинг гормон (ГнРГ) инициирует синтез и выброс лютеинизирующего гормона (ЛГ) и фолликулостимулирующего гормона (ФСГ) из гонадотропных клеток. Половая гормональная обратная связь модулирует его секрецию: в средней фазе менструального цикла эстроген усиливает высвобождение ГнРГ, тогда как в лютеиновой фазе — подавляет.
Кортикотропин-рилизинг гормон (КРГ) активирует кортикотропные клетки гипофиза, вызывая синтез адренокортикотропного гормона (АКТГ). АКТГ стимулирует кору надпочечников к выделению кортизола, который через гипоталамо-гипофизарно-надпочечниковую ось осуществляет отрицательную обратную связь, подавляя высвобождение КРГ.
Гормон роста-рилизинг гормон (ГР-РГ) активирует соматотропные клетки передней доли, вызывая секрецию гормона роста (ГР). При низком уровне ГР и повышенной потребности в анаболических процессах выделение ГР-РГ повышается; однако действие этого гормона ограничивается гормоном роста-ингибирующим гормоном (ГИ-Г), синтезируемым гипоталамусом в ответ на высокий уровень ГР в крови. ГИ-Г связывается с рецепторами соматотропных клеток, снижая их чувствительность к ГР-РГ и образуя классический отрицательный обратный цикл.
Пролактин-ингибиторный гормон (ПИГ) — дофамин — вырабатывается в аркуатных ядрах гипоталамуса и подавляет секрецию пролактина лактотропными клетками передней доли. Уровень дофамина снижается при грудном вскармливании, что позволяет пролактину резко возрасти, тогда как в условиях, не требующих лактации, дофамин активно подавляет пролактин, обеспечивая экономию энергии. Помимо дофамина, гипоталамус синтезирует стимулятор секреции пролактина, образуя двустороннюю регуляцию лактотрофов.
 Рис. 4. Схема гипоталамо‑гипофизарного трактa.
Рис. 4. Схема гипоталамо‑гипофизарного трактa.
Cs4652, CC BY-SA 4.0 https://creativecommons.org/licenses/by-sa/4.0, via Wikimedia Commons
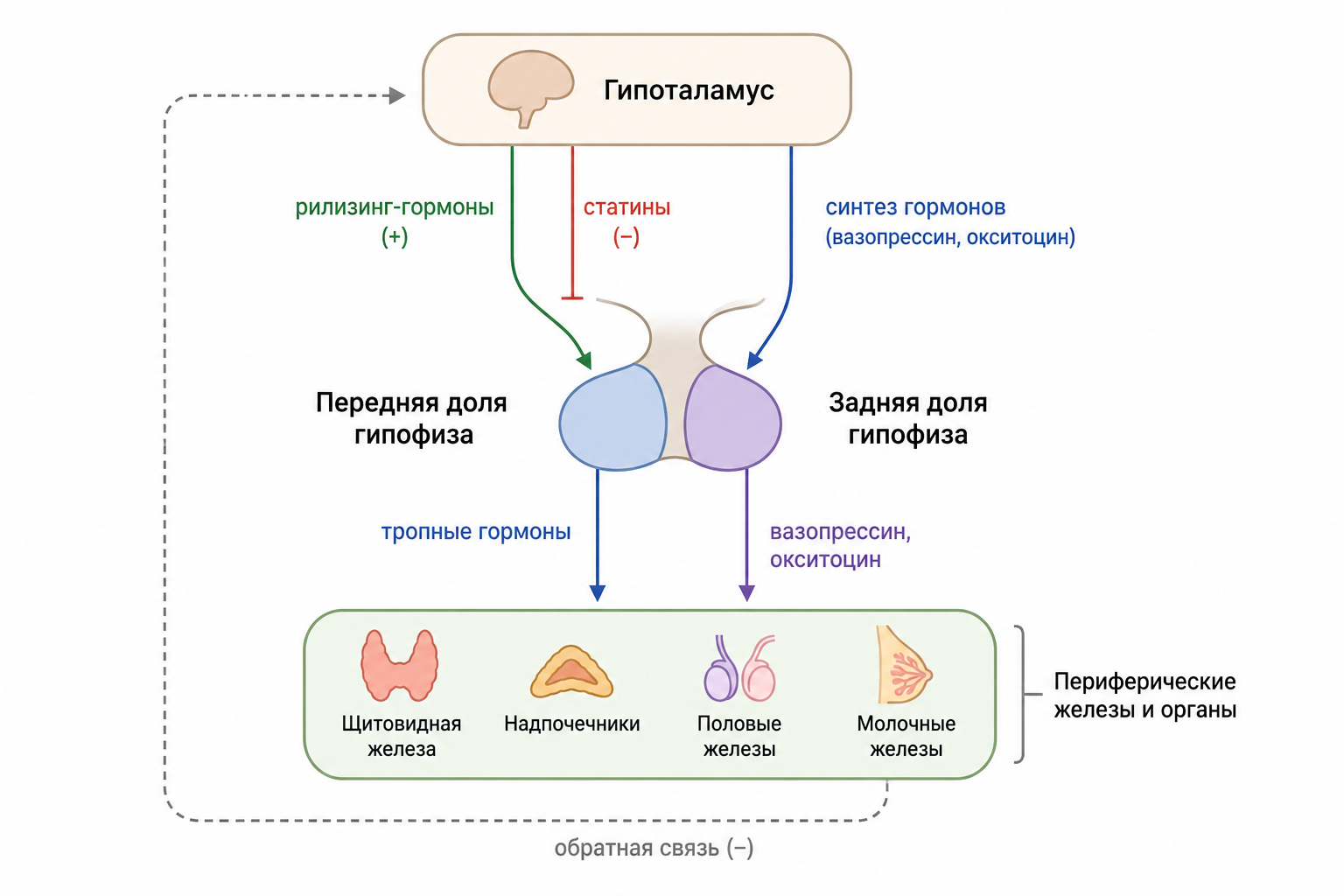 Рис. 5. Схема гипоталамо‑гипофизарного трактa (дополнительная схема).
Рис. 5. Схема гипоталамо‑гипофизарного трактa (дополнительная схема).
Большинство гипоталамических рилизинг- и ингибирующих гормонов синтезируются в нервных окончаниях гипоталамуса; их электростимуляция приводит к последовательному высвобождению в гипофизарный портальный кровоток. Попадая в микро-кровеносные синусоиды передней доли, гормоны взаимодействуют со специфическими рецепторами на разных типах гормон-продуцирующих клеток. Специализированные гипоталамические зоны локализуют нейроны, продуцирующие эти гормоны; тела нейронов отдают начало волокнам срединного возвышения, формирующим гипофизарно-гипоталамический тракт. Гормоны обходят гематоэнцефалический барьер, перемещаясь по портальной системе сосудов, что обеспечивает быстрый и концентрированный эффект, позволяя небольшим изменениям в уровнях гипоталамических гормонов вызывать значительные колебания в секреции гормонов передней доли.
Наличие отрицательной обратной связи характерно для всех пар «гипоталамический гормон — передний гипофиз»: высокий уровень ТТГ подавляет ТРГ, высокий уровень АКТГ (через кортизол) подавляет КРГ, высокий уровень ЛГ/ФСГ подавляет ГнРГ, высокий уровень гормона роста подавляет ГР-РГ через ГИ-Г, а повышенный пролактин подавляет дофаминовые нейроны, уменьшая ПИГ-ингибирование. Эти взаимосвязанные комплексы регуляции гарантируют стабильность эндокринной системы.
Вегетативная и сердечно-сосудистая регуляция
Гипоталамус выступает центральным регулятором вегетативных функций, и его стимуляция способна вызывать разнообразные нейрогенные воздействия на сердечно-сосудистую систему. Активация задней и латеральной областей повышает артериальное давление и частоту сердечных сокращений за счет влияния на центры ретикулярной формации моста и продолговатого мозга, тогда как преоптическая область производит противоположный эффект — снижает как давление, так и ЧСС. Через двусторонние связи с лимбической системой гипоталамус координирует работу автономной нервной системы. Его сигналы направляются к ретикулярной формации ствола мозга (средний мозг, мост, продолговатый мозг), а оттуда к периферическим нервам; одновременно существуют восходящие пути к переднему таламусу и лимбической коре. Главным трактом является нисходящий пучок из большой передней области гипоталамуса, проходящий через его середину и достигающий ретикулярной формации, а также короткие пути между формацией, таламусом и гипоталамусом.
Терморегуляция
Передняя часть гипоталамуса является ключевым центром терморегуляции. В преоптической области расположены температурочувствительные нейроны (термосенсоры), активность которых меняется при изменении температуры протекающей крови. Стимуляция этой зоны запускает механизмы возвращения температуры тела к норме. Хирургические вмешательства в области гипоталамуса почти неизменно вызывают тяжёлую лихорадку, а в редких случаях — гипотермию, демонстрируя двухполярный потенциал его терморегуляторных механизмов. Механическое давление на преоптическую область (например, со стороны опухоли) смещает «заданную величину» (set‑point) вверх, запуская каскад реакций для повышения температуры. Внезапное повышение этого уровня (вследствие деструкции тканей, пирогенных веществ или дегидратации) сопровождается ознобом, ощущением холода, сосудистым спазмом и мышечной дрожью. Когда вызывающий фактор устранён, set‑point возвращается к норме, и при температуре 40 °C гипоталамус воспринимает её как избыточную, включая механизмы теплоотдачи.
Водно-электролитный баланс и регуляция жажды
Регуляция водного баланса обеспечивается супраоптическим ядром гипоталамуса, продуцирующим антидиуретический гормон (АДГ). При повышении осмолярности крови осморецепторы фиксируют сморщивание клеток, что стимулирует секрецию АДГ; концентрация гормона в плазме может измениться всего за несколько минут, а при избытке воды его выделение практически блокируется. Разрушение супраоптического ядра или пересечение нейрогипофиза приводит к несахарному диабету с экскрецией до 10–20 л гипоосмолярной мочи в сутки. Латеральные области гипоталамуса также участвуют в контроле жажды: здесь расположены рецепторы, чувствительные к осмолярности и ангиотензину II, а их поражение вызывает адипсию — полный отказ от питья даже при обезвоживании организма.
Симпатическая и парасимпатическая регуляция
Симпатическая нервная система, находящаяся под контролем гипоталамуса, способна к «массивному разряду» — одновременной активации большинства её звеньев. Такая реакция возникает при сильном страхе или интенсивной боли, вызывая расширение зрачков, учащение сердцебиения, сужение сосудов кожи и быструю мобилизацию ресурсов. В отличие от неё, парасимпатическая система преимущественно управляет отдельными органами, вызывая возбуждение или торможение. Типичные парасимпатические рефлексы включают секрецию пищеварительных соков, дефекацию, мочеиспускание, а у мужчин — эрекцию.
Мочеиспускание и дефекация реализуются как спинальные рефлексы с крестцовой обратной связью, однако гипоталамус оказывает на них модулирующее влияние. Половые рефлексы также осуществляются на крестцовом уровне, но могут инициироваться как психическими стимулами из головного мозга, так и периферическими. В сосудистой системе гипоталамус влияет на сосудодвигательный центр: стимуляция определённых его областей вызывает вазодилатацию или вазоконстрикцию, а дисфункция приводит к нарушениям артериального давления и вазомоторным реакциям.
Энергетический гомеостаз: центры голода и насыщения
В аркуатных ядрах гипоталамуса сосредоточены две противоположные популяции нейронов, образующие центральный узел интеграции периферических сигналов. Проопиомеланокортиновые (РОМС-) нейроны синтезируют α‑меланоцитостимулирующий гормон (α‑MSH) и кокаин- и амфетамин-опосредованные транскрипты (CART); их активация подавляет приём пищи и повышает расход энергии. Противоположную орексигенную ветвь представляют нейроны, продуцирующие нейропептид Y (NPY) и меланин-опосредованный белок (AGRP); их деятельность усиливает потребление пищи и снижает энергетические затраты. Оба подкласса обогащены рецепторами лептина, инсулина, холецистокинина и грелина.
Лептин, секреция которого пропорциональна объёму жировой ткани, проникает через гематоэнцефалический барьер и связывается с рецепторами на РОМС-нейронах. Это подавляет продукцию NPY и AGRP, усиливает секрецию α‑MSH, повышает симпатическую активность и ускоряет метаболизм. Генетические нарушения синтеза лептина или чувствительности его рецепторов сопровождаются гиперфагией и тяжёлым ожирением. Высвобождаемый РОМС-нейронами α‑MSH действует на меланокортиновые рецепторы (MCR‑3 и MCR‑4) в паравентрикулярных ядрах, ограничивая приём пищи. Мутации гена MCR‑4 объясняют 5–6 % случаев раннего тяжёлого детского ожирения, а гиперактивация системы может способствовать анорексии. AGRP, в свою очередь, является естественным inverse agonist для MCR-3 и MCR-4, конкурируя с α‑MSH и усиливая пищевой приём синергетически с NPY.
Боковой гипоталамус играет центральную роль в поддержании базовых мотивационных состояний: его двустороннее поражение приводит к отказу от воды и пищи и выраженной пассивности, а стимуляция вызывает жажду, голод, повышение двигательной активности и агрессию. Вентромедиальные ядра функционируют антагонистически: их поражение вызывает безудержное потребление пищи и гиперактивность, а стимуляция — чувство насыщения и успокоение, подтверждая роль этих структур как центра насыщения.
Гастроинтестинальные гормоны заполняют промежуточное звено между приёмом пищи и центральным регулятором голода. Грелин, выделяемый клетками желудка, достигает пика в состоянии голода и стимулирует орексигенные нейроны. Противоположный сигнал дают гормоны сытости: холецистокинин, пептид YY, глюкагоноподобный пептид и инсулин. Сенсорные рецепторы ротовой полости ограничивают объём съеденного за счёт механических и вкусовых ощущений на 20–40 минут, после чего более сильный сигнал поступает от растяжения ЖКТ. К анорексигенным факторам также относятся серотонин, норадреналин, кортикотропин-рилизинг гормон; к орексигенным — меланин-концентрирующий гормон, орексины А и В, эндорфины, галанин, глутамат, ГАМК и кортизол. Терморегулятивные потребности также модулируют сети голода: при низкой температуре ускоряется накопление жировых запасов.
Репродуктивная функция и роль окситоцина
Гонадотропин-рилизинг гормон (ГнРГ) представляет собой пептидную цепочку из 10 аминокислот, которая стимулирует секрецию лютеинизирующего гормона (ЛГ) и фолликулостимулирующего гормона (ФСГ), контролируя стероидогенез и гаметогенез. У некоторых видов животных шишковидная железа (эпифиз) подавляет продукцию гонадотропных гормонов в зимние месяцы, однако через 4 месяца бездеятельности гонады освобождаются из-под тормозных влияний, достигая максимальной активности весной. У человека роль эпифиза окончательно не установлена, но опухоли в этой области приводят к гипер- или гипогонадизму. Регуляция женского месячного цикла осуществляется по принципу отрицательной обратной связи: колебания активности в системе «гипоталамус — передняя доля гипофиза — яичники» контролируют продукцию эстрадиола и прогестерона. Стимуляция самых передних и задних отделов гипоталамуса связана с сексуальным возбуждением.
Окситоцин является мощным стимулятором сокращений матки в конце гестации и рассматривается как один из возможных механизмов наступления родов. Кроме того, он играет ключевую роль в лактации: сосание ребёнком соска стимулирует нейроны паравентрикулярного и супраоптического ядер, окситоцин выделяется в заднюю долю гипофиза и с током крови вызывает сокращение миоэпителиальных клеток молочных желез, обеспечивая выделение молока менее чем через минуту после начала сосания. Секреция пролактина лактотропными клетками регулируется двусторонне: подавляется пролактин-ингибирующим гормоном (ПИГ) — дофамином (синтезируется в аркуатных ядрах), и стимулируется специфическими гипоталамическими факторами. Во время грудного вскармливания уровень дофамина снижается, позволяя пролактину расти.
Стимуляция перивентрикулярных ядер, прилегающих к третьему желудочку, и центрального серого вещества среднего мозга провоцирует реакции страха и наказания, формируя избегающее поведение в рамках системы отрицательного подкрепления.
Заключение
Гипоталамус является важнейшим интегративным центром, объединяющим нервную и эндокринную регуляцию. Через сложную систему ядер и нейропептидов он контролирует жизненно важные функции: от терморегуляции и водно-солевого баланса до пищевого поведения, энергетического гомеостаза и репродукции. Центры голода и насыщения, расположенные соответственно в латеральном и вентромедиальном гипоталамусе, тонко настраиваются периферическими гормонами, такими как лептин и грелин, а дисфункция этих структур приводит к тяжёлым метаболическим расстройствам. Регуляция водного баланса посредством антидиуретического гормона предотвращает критические потери жидкости, а взаимодействие рилизинг- и ингибирующих гормонов с гипофизом гарантирует стабильность всей эндокринной системы. Управляя вегетативными рефлексами, сердечно-сосудистой деятельностью и эмоциональными реакциями, гипоталамус обеспечивает непрерывную адаптацию организма к изменяющимся условиям внешней и внутренней среды, оставаясь незаменимым дирижёром нашего физиологического благополучия.