Механизмы генерации и проведения электрических сигналов в нейроне
Нервная система работает на электричестве, но это электричество особого рода: оно рождается не в проводах, а в тончайшем слое заряженных частиц, выстроившихся вдоль клеточной мембраны. Чтобы понять, как нейрон генерирует, проводит и передаёт сигнал, нужно разобраться в нескольких базовых механизмах. В этой статье мы последовательно пройдём пять этапов: от физико-химических основ мембранных потенциалов к формированию потенциала покоя, далее к потенциалу действия, проведению импульса по аксону и, наконец, к синаптической передаче и интеграции сигналов. Каждый раздел построен так, чтобы предыдущий становился фундаментом для следующего.
1. Физико-химические основы: ионные градиенты и математические модели
Мембранные потенциалы существуют практически у всех клеток организма, но именно нейроны и мышечные клетки используют их для быстрой передачи сигналов. В основе этого явления лежат три фактора: разная концентрация ионов внутри и снаружи клетки, избирательная проницаемость мембраны и работа ионных насосов. Эти три составляющих неразрывны: убрать любую из них — и электрический сигнал исчезнет.
1.1. Разделение зарядов и электрический дипольный слой
Важно понимать, что для создания мембранного потенциала (например, −70 мВ) не требуется перемещения огромного количества ионов. На самом деле перемещается лишь ничтожно малая часть — от 1/3 000 000 до 1/100 000 000 от общего числа положительных ионов внутри клетки. Эти ионы выстраиваются непосредственно вдоль мембраны, образуя электрический дипольный слой: изнутри скапливаются отрицательные заряды, снаружи — положительные. В глубине цитоплазмы и во внеклеточной жидкости количество положительных и отрицательных зарядов остаётся равным — там сохраняется состояние электронейтральности.
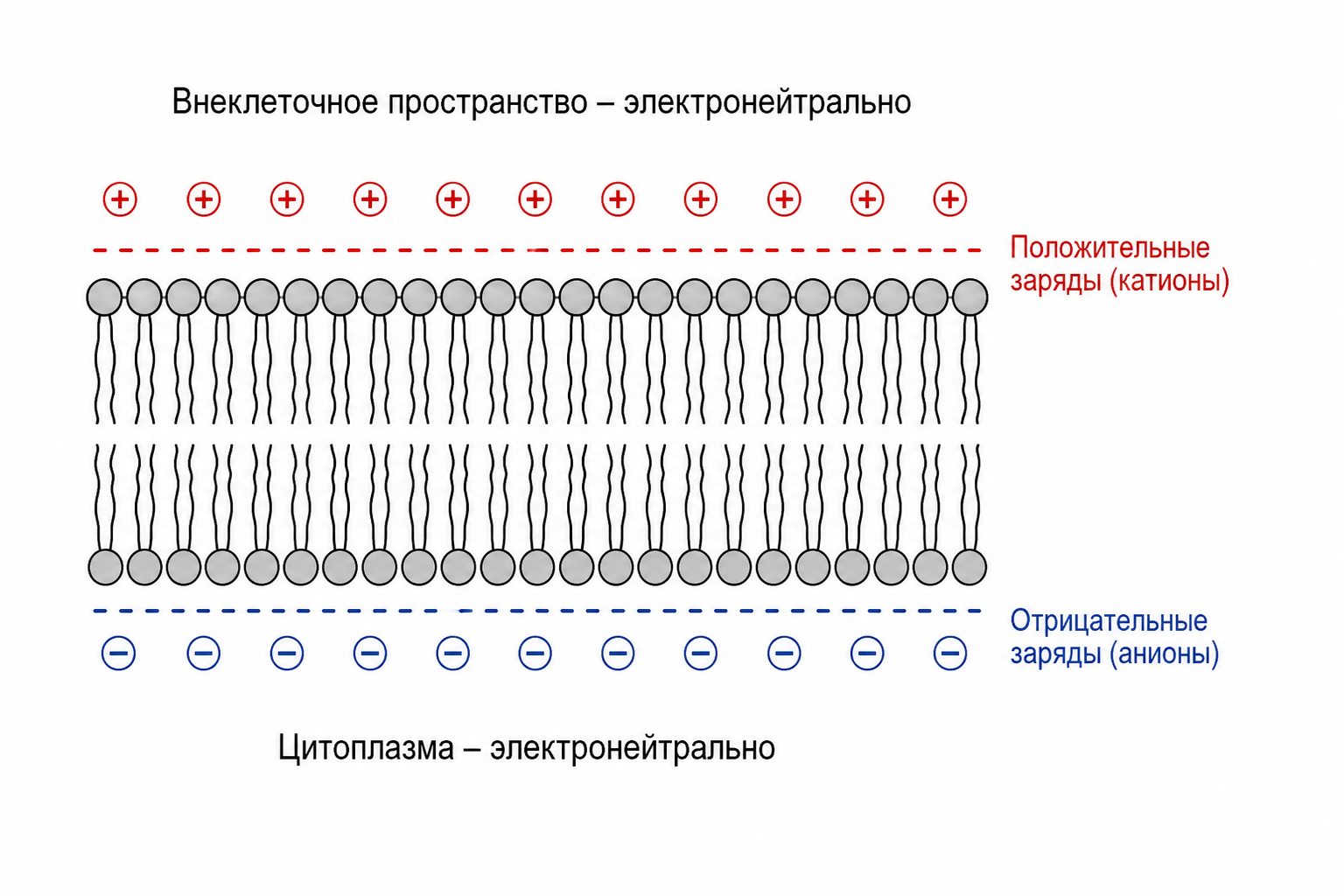 Рис. 1. Схема электрического дипольного слоя мембраны: отрицательные заряды (анионы) вдоль внутренней поверхности, положительные заряды (катионы) вдоль наружной; в глубине цитоплазмы и снаружи — электронейтральность.
Рис. 1. Схема электрического дипольного слоя мембраны: отрицательные заряды (анионы) вдоль внутренней поверхности, положительные заряды (катионы) вдоль наружной; в глубине цитоплазмы и снаружи — электронейтральность.
1.2. Основные ионные градиенты
Для генерации потенциалов в нейронах млекопитающих критически важны три иона, плюс одна группа крупных анионов, которые не проходят через мембрану. Их концентрации поддерживаются в строго определённом соотношении.
1. Калий (K⁺) — главный внутриклеточный катион. Концентрация внутри — 140 мэкв/л, снаружи — 4 мэкв/л. Стремится выйти из клетки.
2. Натрий (Na⁺) — главный внеклеточный катион. Концентрация внутри — 14 мэкв/л, снаружи — 142 мэкв/л. Стремится войти в клетку.
3. Хлор (Cl⁻) — его концентрация выше снаружи (103 мэкв/л) и низка внутри (8 мэкв/л).
4. Анионы белков и фосфатов — находятся внутри клетки в высокой концентрации и не способны диффундировать через мембрану, обеспечивая базовую электроотрицательность внутренней среды.
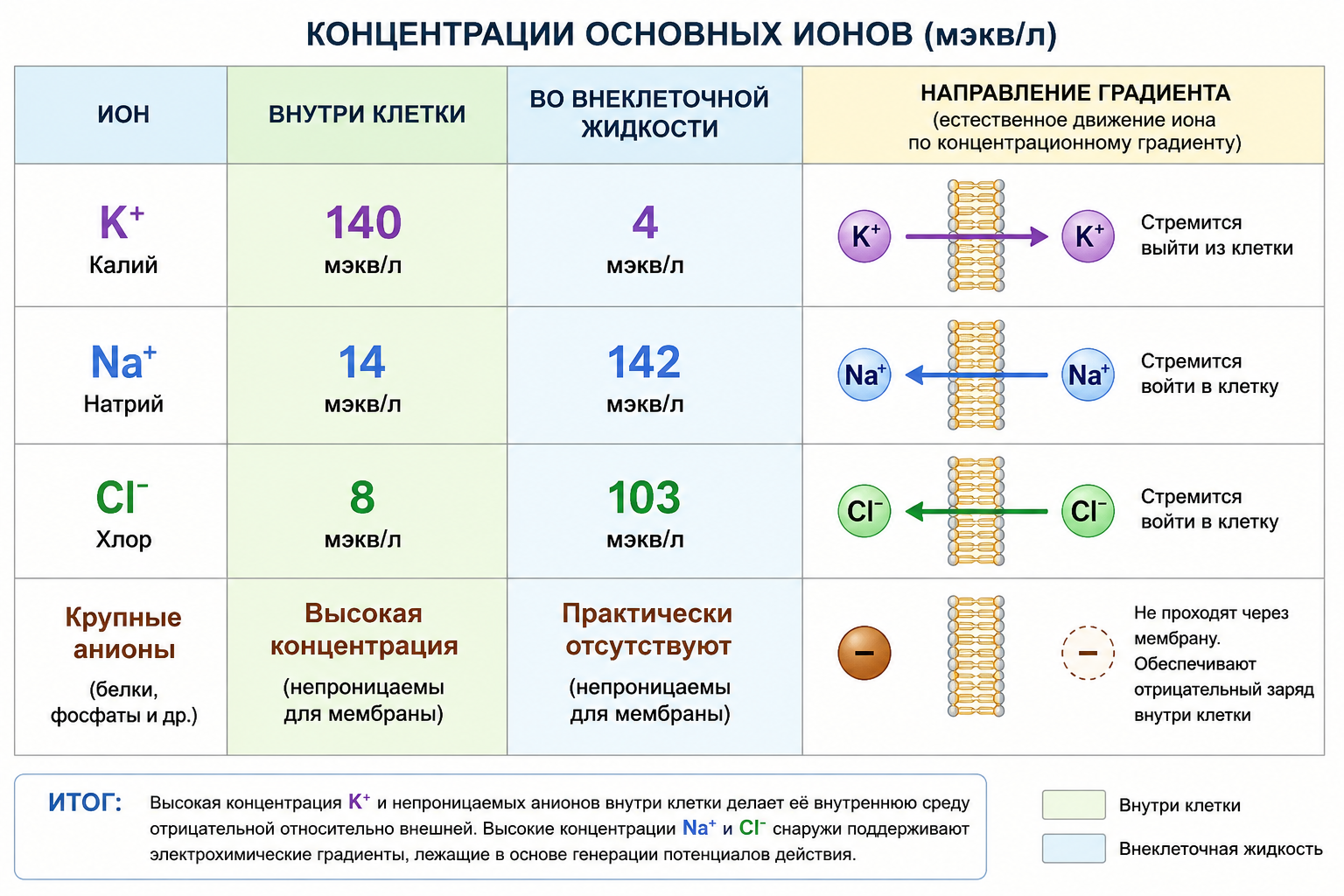 Рис. 2. Сравнительная таблица (или столбчатая диаграмма) концентраций K⁺, Na⁺, Cl⁻ и крупных анионов внутри клетки и во внеклеточной жидкости, с указанием стрелками направления градиента для каждого иона.
Рис. 2. Сравнительная таблица (или столбчатая диаграмма) концентраций K⁺, Na⁺, Cl⁻ и крупных анионов внутри клетки и во внеклеточной жидкости, с указанием стрелками направления градиента для каждого иона.
1.3. Уравнение Нернста: потенциал равновесия
Если мембрана проницаема только для одного иона, возникает потенциал Нернста — это такая разность потенциалов, которая точно уравновешивает стремление иона диффундировать по градиенту концентрации. Иначе говоря, в этой точке электрическая сила, тянущая ион обратно, становится равной диффузионной силе, толкающей его по градиенту. Для одновалентных ионов при температуре тела (37 °C) формула выглядит так:
ЭДС (мВ) = ± 61 × log(Cᵢ / Cₒ)
Здесь Cᵢ — концентрация иона внутри клетки, Cₒ — снаружи. Для калия (K⁺) потенциал Нернста составляет −94 мВ (отрицательный заряд внутри). Для натрия (Na⁺) он равен +61 мВ (положительный заряд внутри). Знак определяется тем, какой ион мы рассматриваем и в какую сторону направлен его градиент.
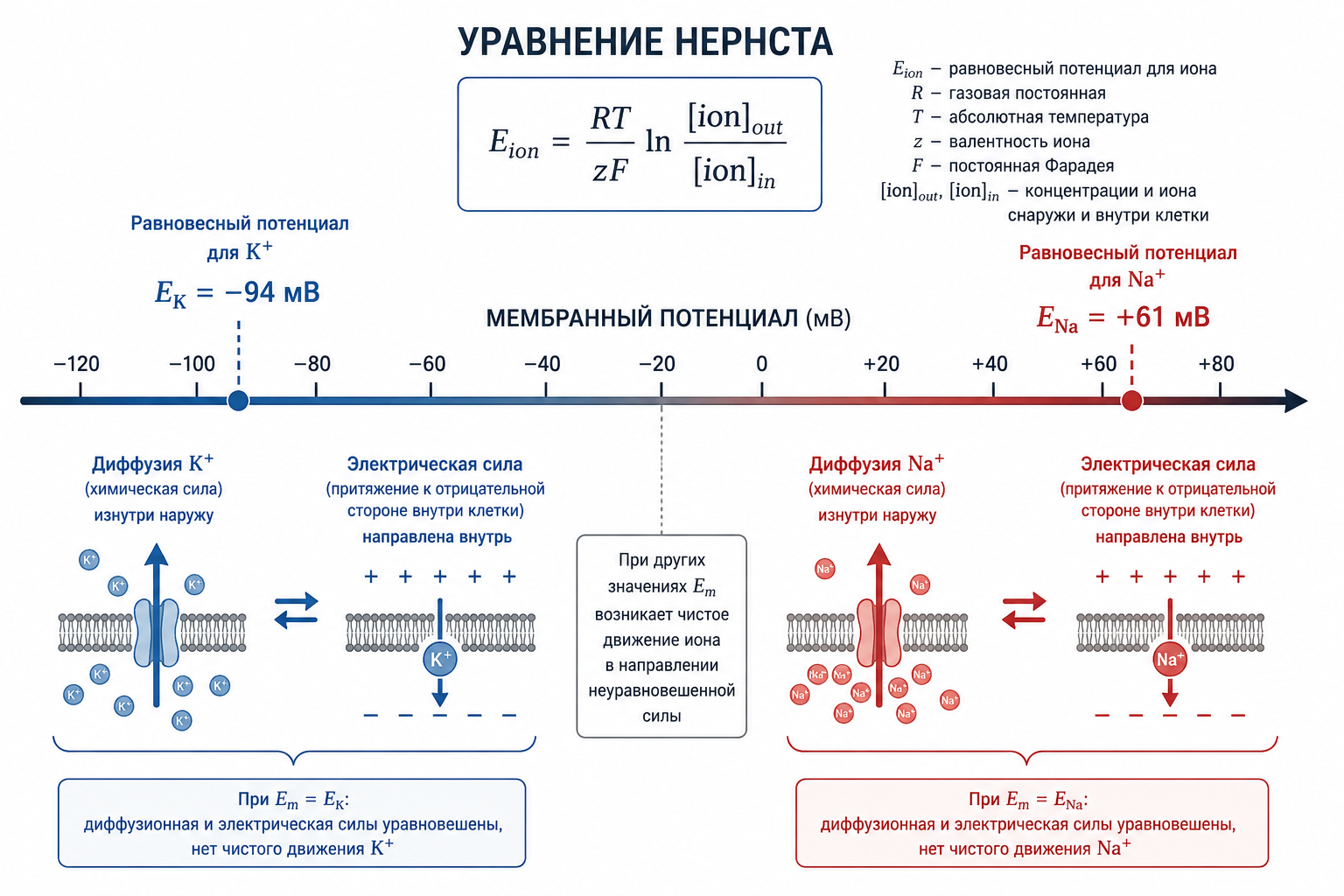 Рис. 3. Графическая иллюстрация уравнения Нернста: на одной оси — мембранный потенциал, показаны равновесные значения для K⁺ (−94 мВ) и Na⁺ (+61 мВ) с указанием направления диффузии и уравновешивающей электрической силы.
Рис. 3. Графическая иллюстрация уравнения Нернста: на одной оси — мембранный потенциал, показаны равновесные значения для K⁺ (−94 мВ) и Na⁺ (+61 мВ) с указанием направления диффузии и уравновешивающей электрической силы.
1.4. Уравнение Гольдмана (Гольдмана–Ходжкина–Катца)
В реальности мембрана проницаема для нескольких ионов одновременно. Суммарный мембранный потенциал рассчитывается по уравнению Гольдмана, которое учитывает три фактора: полярность электрического заряда каждого иона, проницаемость (P) мембраны для каждого иона и концентрации (C) соответствующих ионов внутри (i) и снаружи (o).
Ключевой вывод уравнения: степень вклада иона в величину мембранного потенциала прямо пропорциональна проницаемости мембраны для него. Если проницаемость для натрия и хлора равна нулю, то мембранный потенциал будет равен потенциалу Нернста для калия.
Именно быстрые изменения проницаемости для натрия и калия являются механизмом передачи сигналов (потенциала действия), в то время как проницаемость для хлора меняется незначительно. Это разграничение очень важно: хлор скорее выполняет функцию «стабилизатора», тогда как натрий и калий — рабочие ионы сигнала.
2. Потенциал покоя: механизмы и вклад различных факторов
Потенциал покоя крупных нервных волокон в норме составляет около −90 мВ (в соме нейрона он может быть выше — около −65…−70 мВ). Это означает, что внутренняя поверхность мембраны на соответствующее количество милливольт отрицательнее внеклеточной жидкости. Цифра −90 мВ — не случайное значение, а результат математически выверенного баланса трёх факторов, которые мы разберём по очереди.
2.1. Вклад диффузии калия через каналы утечки
Главный вклад в потенциал покоя вносит калий. В мембране нейрона есть особые белки — калиевые каналы утечки (каналы с двумя тандемными поровыми доменами). В состоянии покоя они остаются открытыми, позволяя ионам калия постоянно «вытекать» из клетки по градиенту концентрации. Мембрана в покое примерно в 100 раз более проницаема для K⁺, чем для Na⁺.
Если бы потенциал покоя зависел только от калия, он составил бы −94 мВ (потенциал Нернста для K⁺). Однако из-за того, что мембрана всё же пропускает мизерное количество натрия внутрь, суммарный потенциал диффузии смещается до −86 мВ. То есть даже небольшой «подток» натрия ощутимо сдвигает потенциал в положительную сторону.
2.2. Вклад натрий-калиевого насоса
Насос не просто поддерживает градиенты, он сам по себе является электрогенным — то есть создаёт электрический ток. За один цикл работы насос выкачивает 3 иона натрия (Na⁺) наружу и закачивает 2 иона калия (K⁺) внутрь. Поскольку наружу выносится на один положительный заряд больше, чем заносится внутрь, создаётся постоянный дефицит положительных ионов в клетке. Это добавляет ещё −4 мВ к потенциалу покоя.
Итог: −86 мВ (диффузия) + (−4 мВ) (насос) = −90 мВ. Таким образом, потенциал покоя складывается из двух источников — пассивной диффузии калия и активной электрогенной работы насоса.
 Рис. 4. Схема работы Na⁺/K⁺-насоса: за один цикл 3 Na⁺ выносятся наружу и 2 K⁺ заносятся внутрь; показано расщепление АТФ. Также показаны натриевые и калиевые каналы утечки (синим и розовым соответственно).
Рис. 4. Схема работы Na⁺/K⁺-насоса: за один цикл 3 Na⁺ выносятся наружу и 2 K⁺ заносятся внутрь; показано расщепление АТФ. Также показаны натриевые и калиевые каналы утечки (синим и розовым соответственно).
Klkelly02, CC BY-SA 4.0 https://creativecommons.org/licenses/by-sa/4.0, via Wikimedia Commons
2.3. Роль недиффундирующих анионов
Внутри нейрона находятся крупные отрицательно заряженные частицы — анионы белков, фосфатов и сульфатов. Они слишком велики, чтобы пройти через ионные каналы. Когда калий стремится выйти из клетки по градиенту, эти анионы остаются внутри, «притягивая» положительные заряды обратно и обеспечивая базовую электроотрицательность цитоплазмы. Их роль — удерживать клетку в состоянии «отрицательного баланса» даже тогда, когда калий пытается её покинуть.
2.4. Физика процесса: электрический дипольный слой
Важно понимать, что заряд распределён не по всей клетке равномерно. Отрицательные заряды (анионы) выстраиваются вдоль внутренней поверхности мембраны, а положительные заряды (катионы) — вдоль наружной. Этот тонкий слой зарядов непосредственно на границе раздела сред и создаёт измеряемое напряжение. В глубине самого нейрона (в цитоплазме) среда остаётся электронейтральной. Именно поэтому потенциал в −90 мВ существует в очень узком слое толщиной в несколько нанометров, а не «размазан» по всему объёму клетки.
3. Потенциал действия: молекулярные ворота и фазовый цикл
Потенциал действия — это быстрый, самораспространяющийся цикл изменений мембранного потенциала, который служит основным сигналом в нервной системе. Он работает по принципу «всё или ничего»: импульс либо не возникает вовсе (если стимул подпороговый), либо имеет максимальную и стандартную амплитуду при достижении порога. Промежуточных, «половинчатых» импульсов не бывает.
3.1. Пороговый потенциал и положительная обратная связь
Для возникновения потенциала действия необходима деполяризация мембраны от уровня покоя (−70…−90 мВ) до порога возбуждения (обычно около −55 мВ). Когда мембранный потенциал достигает порога, открывается критическое количество потенциал-зависимых Na⁺-каналов. Вход ионов Na⁺ дополнительно деполяризует мембрану, что приводит к открытию ещё большего числа натриевых каналов. Этот лавинообразный процесс (цикл Ходжкина) обеспечивает стремительный взлёт потенциала — то, что и составляет фазу деполяризации.
3.2. Работа ворот натриевого канала
Потенциал-зависимый Na⁺-канал нейрона — это сложный белковый комплекс, имеющий две системы ворот, которые работают в разном временном ритме. Их скоординированная работа и обеспечивает характерную форму потенциала действия.
Активационные ворота (m-ворота). Находятся снаружи канала. В покое они закрыты. При деполяризации открываются мгновенно, обеспечивая фазу деполяризации.
Инактивационные ворота (h-ворота). Находятся внутри канала. В покое они открыты («взведены»). Деполяризация заставляет их закрыться, но они реагируют медленнее активационных — с задержкой в доли миллисекунды.
Можно выделить три состояния канала:
Закрыт (в покое): активационные ворота закрыты, инактивационные — открыты. Канал готов к работе.
Открыт (пик деполяризации): обе системы ворот открыты одновременно на кратчайший миг.
Инактивирован (начало реполяризации): активационные ворота ещё могут быть открыты, но инактивационные («шарик на верёвочке») уже закупорили пору изнутри. В этом состоянии канал не может открыться повторно.
 Рис. 5. Схема трёх состояний натриевого канала: 1) покой (m-закрыта, h-открыта); 2) открытое состояние (m-открыта, h-открыта); 3) инактивация (m-открыта, h-закрыта).
Рис. 5. Схема трёх состояний натриевого канала: 1) покой (m-закрыта, h-открыта); 2) открытое состояние (m-открыта, h-открыта); 3) инактивация (m-открыта, h-закрыта).
Fotfoti, CC BY-SA 4.0 https://creativecommons.org/licenses/by-sa/4.0, via Wikimedia Commons
3.3. Фазы потенциала действия
Деполяризация (Upstroke). Массивный вход Na⁺ устремляет потенциал к его равновесному значению (+61 мВ). В крупных волокнах потенциал проскакивает нулевой уровень (овершут), достигая +30…+35 мВ.
Реполяризация (Downstroke). На пике потенциала действия натриевые каналы инактивируются, а потенциал-зависимые калиевые каналы открываются с задержкой. Ионы K⁺ выходят из клетки, возвращая мембрану к отрицательному заряду.
Следовая гиперполяризация (Undershoot). Калиевые каналы закрываются медленно, и на короткое время мембранный потенциал становится даже более отрицательным, чем в покое, приближаясь к потенциалу Нернста для калия (−94 мВ).
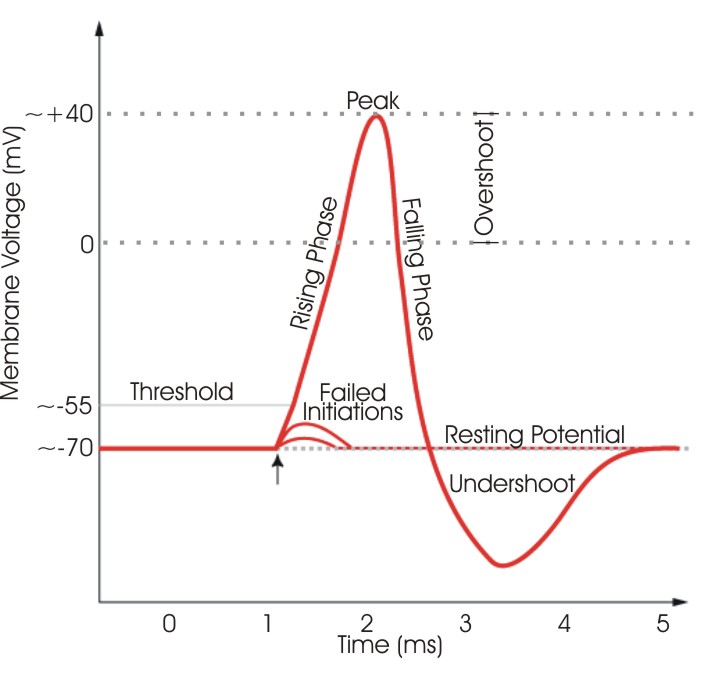 Рис. 6. График потенциала действия с обозначением всех фаз: деполяризация, овершут (+30…+35 мВ), реполяризация, следовая гиперполяризация.
Рис. 6. График потенциала действия с обозначением всех фаз: деполяризация, овершут (+30…+35 мВ), реполяризация, следовая гиперполяризация.
The original uploader was Synaptidude at English Wikipedia., GFDL 1.2 http://www.gnu.org/licenses/old-licenses/fdl-1.2.html, via Wikimedia Commons
3.4. Молекулярный сенсор напряжения
Чувствительность каналов к электричеству обеспечивается спиралью S4 в составе белка. Она содержит положительно заряженные аминокислоты (аргинин, лизин) через каждые три позиции. При деполяризации (когда внутри становится «плюс») эти заряды отталкиваются наружу, вызывая поворот или смещение спирали, что механически открывает ворота канала. Иными словами, канал буквально «чувствует» электрическое поле и преобразует его в механическое движение своих внутренних частей.
3.5. Роль кальция в возбудимости
Ионы Ca²⁺ действуют как «стабилизаторы» натриевых каналов, связываясь с их внешней стороной и повышая порог активации. При гипокальциемии (дефиците кальция) натриевые каналы открываются при гораздо меньшем сдвиге потенциала. Это приводит к спонтанному возбуждению нервов и судорогам (тетании). На этом примере хорошо видно, что электрические свойства нейрона зависят не только от натрия и калия, но и от тонкой химической «настройки» мембраны.
4. Рефрактерность и проведение импульса: биофизический аспект
После генерации потенциала действия мембрана на некоторое время теряет свою возбудимость. Это критически важно для того, чтобы импульс не «эховал» назад и не сливался с последующими сигналами. Существуют два типа рефрактерности, и различать их необходимо для понимания частотных свойств нейрона.
4.1. Рефрактерный период (период невозбудимости)
Абсолютный рефрактерный период (АРП) перекрывает почти всю длительность потенциала действия. Его причина — закрытые инактивационные ворота (h-ворота) натриевых каналов. Пока мембрана не реполяризуется хотя бы до порогового уровня, «шарик» не выйдет из поры, и никакой стимул не откроет канал снова. Смысл АРП — ограничить максимальную частоту разрядов (для крупных волокон — до 2500 имп/с) и гарантировать одностороннее движение импульса.
Относительный рефрактерный период (ОРП) следует сразу за АРП и совпадает с фазой следовой гиперполяризации. Натриевые каналы уже начали восстанавливаться, но калиевая проводимость всё ещё аномально высока. В этот момент потенциал действия можно вызвать, но требуется сверхпороговый стимул, так как мембрана находится дальше от порога, чем в покое.
4.2. Проведение импульса: локальные токи
Потенциал действия — это не движение ионов вдоль аксона, а последовательная перезарядка соседних участков мембраны. В месте потенциала действия внутренняя сторона мембраны становится положительной. Возникают локальные круговые токи: положительные заряды внутри аксона перетекают к соседним отрицательно заряженным участкам покоя, деполяризуя их до порога и вызывая там новый потенциал действия. Так сигнал шаг за шагом «перепрыгивает» вдоль волокна.
4.3. Кабельные свойства нейрона
Эффективность проведения зависит от пассивных электрических характеристик мембраны. Постоянная времени (τ) — это время, за которое мембрана «заряжается». Чем меньше τ, тем быстрее мембрана деполяризуется до порога в ответ на ток. Постоянная длины (λ) — расстояние, на которое ток распространяется пассивно, прежде чем затухнуть на 63%.
Постоянная длины λ увеличивается при росте сопротивления мембраны (Rₘ) — то есть когда меньше утечек тока — и уменьшении внутреннего сопротивления цитоплазмы (Rᵢ), то есть при более широком диаметре аксона. Чем больше λ, тем на большее расстояние «забрасываются» локальные токи и тем выше скорость проведения. Эти соотношения часто становятся камнем преткновения, но их логика проста: чем «лучше изолирована» мембрана и чем «толще кабель», тем дальше сигнал проходит без затухания.
4.4. Сальтаторное (скачкообразное) проведение
У позвоночных для радикального ускорения проведения используется миелин. В периферической нервной системе его формируют шванновские клетки, в центральной нервной системе — олигодендроциты. Миелин увеличивает сопротивление мембраны (Rₘ), то есть ток не вытекает наружу, и снижает электрическую ёмкость мембраны (Cₘ) в 50 раз. Перехваты Ранвье — единственные места, где миелин прерывается — становятся зонами концентрации почти всех Na⁺-каналов: их плотность здесь достигает 10 000 на мкм² против 20 в безмиелиновых волокнах.
Ток практически мгновенно проходит через изолированный междоузловой сегмент от одного перехвата к другому. Отсюда два главных преимущества сальтаторного проведения:
Скорость. Повышается в 5–50 раз — до 100–120 м/с.
Энергоэффективность. Деполяризуется лишь малая площадь мембраны, поэтому Na⁺/K⁺-насосу нужно перекачивать в 100 раз меньше ионов для восстановления градиента.
4.5. Фактор надёжности
В миелинизированных волокнах потенциал, приходящий от одного узла к другому, обычно в 5 раз выше, чем требуется для достижения порога. Это гарантирует, что сигнал не затухнет при небольших повреждениях или ветвлении аксона. Запас прочности — важное свойство нервной системы: она спроектирована так, чтобы сигнал доходил до адресата даже в неидеальных условиях.
5. Синаптическая передача и интеграция сигналов
Хотя внутри нейрона информация передаётся электрическим путём, в подавляющем большинстве контактов (синапсов) между клетками этот способ сменяется химическим. Это позволяет нервной системе не просто проводить сигнал, но и гибко его обрабатывать: усиливать, тормозить, фильтровать и комбинировать.
5.1. Механизм химической передачи: мост между клетками
Когда потенциал действия достигает окончания аксона (пресинаптической терминали), запускается каскад событий. Деполяризация терминали открывает потенциал-зависимые Ca²⁺-каналы. Концентрация кальция снаружи в 1000 раз выше, чем внутри, поэтому ионы стремительно входят в клетку. Ионы Ca²⁺ связываются с белками (например, синаптотагмином), что заставляет везикулы, наполненные нейромедиатором, сливаться с мембраной и выбрасывать содержимое в синаптическую щель шириной 20–30 нм. Медиатор за доли миллисекунды диффундирует через щель и связывается с белками-рецепторами на мембране следующей клетки.
5.2. Постсинаптические потенциалы: ВПСП и ТПСП
Ответ нейрона зависит не от самого медиатора, а от того, какой канал открывает рецептор. Один и тот же медиатор в разных синапсах может быть как возбуждающим, так и тормозным — важна не молекула, а рецептор и канал, который он открывает.
Возбуждающий постсинаптический потенциал (ВПСП) возникает при открытии каналов для Na⁺ (реже Ca²⁺). Вход положительных зарядов вызывает локальную деполяризацию, которая стремится приблизить мембранный потенциал к порогу возбуждения.
Тормозной постсинаптический потенциал (ТПСП) возникает при открытии каналов для Cl⁻ (вход) или K⁺ (выход). Это вызывает гиперполяризацию, делая внутреннюю сторону мембраны ещё более отрицательной и удаляя потенциал от порога.
5.3. Интеграция и суммация: «вычислительный центр» нейрона
Одиночный ВПСП обычно составляет всего 0,5–1 мВ, что слишком мало для запуска потенциала действия: порог требует сдвига на 15–20 мВ. Поэтому нейрон суммирует все поступающие входы. Это происходит двумя способами.
Временная суммация — сложение эффектов от быстро следующих друг за другом разрядов одной и той же пресинаптической терминали. Если второй стимул приходит до того, как затух первый ВПСП, они накладываются друг на друга.
Пространственная суммация — сложение эффектов от множества разных синапсов, расположенных в разных точках дендритов и сомы, активируемых одновременно.
5.4. Аксонный холмик: место принятия решения
Суммарное изменение потенциала распространяется по телу нейрона (соме). Ключевым участком является начальный сегмент аксона — аксонный холмик. Здесь плотность потенциал-зависимых Na⁺-каналов в 7 раз выше, чем в соме, поэтому порог возбуждения в этой зоне минимален. Если итоговая деполяризация в этой точке достигает порога (около −55 мВ), в аксоне рождается новый потенциал действия, который неослабевающим импульсом бежит к следующей клетке. Таким образом, нейрон выполняет роль аналогового «вычислителя»: он складывает тысячи входных сигналов и принимает бинарное решение — выдать импульс или нет.
Заключение
Мы прошли весь путь электрического сигнала в нервной системе — от ионных градиентов, поддерживаемых на границе мембраны, до бинарного решения, принимаемого аксонным холмиком. На каждом этапе работают одни и те же базовые принципы: разница концентраций ионов, избирательная проницаемость мембраны и активная работа ионных насосов. Эти принципы создают потенциал покоя, который при достижении порога превращается в потенциал действия; потенциал действия распространяется по аксону за счёт локальных токов и (у позвоночных) сальтаторного проведения; а на синапсе электрический сигнал преобразуется в химический, чтобы следующая клетка могла его обработать и принять собственное решение. Понимание этой цепочки — необходимый фундамент для изучения всего, что происходит в нервной системе дальше: от рефлексов до обучения и памяти.