Метаморфозы
Метаморфозы (от греч. metamorphosis — превращение, преобразование) представляют собой видоизменения органов, возникающие, когда дополнительные функции начинают преобладать над главными, либо когда орган в процессе развития приобретает новые задачи. Эти изменения затрагивают как морфологию, так и анатомию растения, являясь следствием его приспособления к особым условиям существования. Термин был введён Карлом Линнеем, который в 1755 г. опубликовал труд «Metamorphosis plantarum». Учение имеет глубокие корни: ещё в XVI веке А. Цезальпин отмечал возможность видоизменения вегетативных органов, считая лепестки изменёнными листьями, а М. Мальпиги рассматривал корень как часть стебля, ветвящегося до волосков. Линней представлял цветок как совокупность одновременно появляющихся листьев, а К. Ф. Вольф свёл все части растения к двум органам — стеблю и листу. Наиболее существенный вклад в развитие учения внёс И. В. Гёте. Он полагал идентичность всех частей растения и вывел их из бокового (аппендикулярного) органа, названного им листом, включая специальные формы, не уподобляя их обычному вегетативному листу (филлу).
Вегетативные органы у растений по своей природе мультифункциональны. Лист, помимо основной фотосинтетической функции, участвует в дыхании и транспирации, может запасать воду, а благодаря железистым волоскам и шипикам защищает организм от поедания. Корень, будучи главным элементом поглощения воды и закрепления в субстрате, одновременно участвует в накоплении запасных веществ. Стебель обеспечивает транспорт веществ между листьями и корнями, удерживает листву и задаёт ориентацию растения. Кроме того, у травянистых и молодых древесных видов стебель способен к фотосинтезу и хранению запасных продуктов.
Классификация и функциональные типы метаморфозов
Функциональные метаморфозы разнообразны и подразделяются на несколько типов. Запасающие метаморфозы (корнеплоды, корневые шишки, клубни, корневища, луковицы, воздушные корни эпифитных растений и пневматофоры) накапливают органические вещества, воду и воздух; они обычно мясистые и богаты паренхимой. Питающие метаморфозы делятся на фотосинтезирующие (кладодии, филлокладии, ассимилирующие корни водных растений и воздушные корни эпифитных орхидей) и органы, поглощающие готовые питательные вещества из тканей других растений (гаустории) или животных (ловчие листья хищных растений). Защитные метаморфозы представляют собой разнообразные колючки, которые следует отличать от шипов-эмергенций, не являющихся метаморфозами вегетативных органов, а лишь их поверхностными разрастаниями. Опорные метаморфозы позволяют растению удерживаться на опоре и обеспечивают ориентацию: к ним относятся корни-прицепки, корни-подпорки, ходульные и досковидные корни, а также усики.
Органы для вегетативного размножения (клубни, корневища, луковицы, столоны) демонстрируют, что метаморфозы, как и обычные вегетативные органы, способны выполнять несколько взаимосвязанных функций. При этом одну и ту же функцию могут выполнять метаморфозы разного происхождения, обладающие как морфологическим, так и анатомическим сходством. Ассимилирующие органы обычно плоские, с хорошо развитой хлоренхимой, расположенной под прозрачной покровной тканью, тогда как органы, исполняющие механическую роль, характеризуются сильным одревеснением, придающим им повышенную твёрдость и прочность.
Для определения того, какой вегетативный орган подвергся видоизменению, применяют три главных критерия: общий план строения, расположение относительно других органов и особенности заложения и развития. Гомологичными называют органы одинакового происхождения, но специализировавшиеся в разных функциях. К таковым относятся корневища, клубни, луковицы, колючки боярышника, кладодии, филлокладии, усики винограда и тыквы. Все они исторически развились из листостебельного побега, что подтверждается их развитием из почек, закладывающихся на конусе нарастания главного побега.
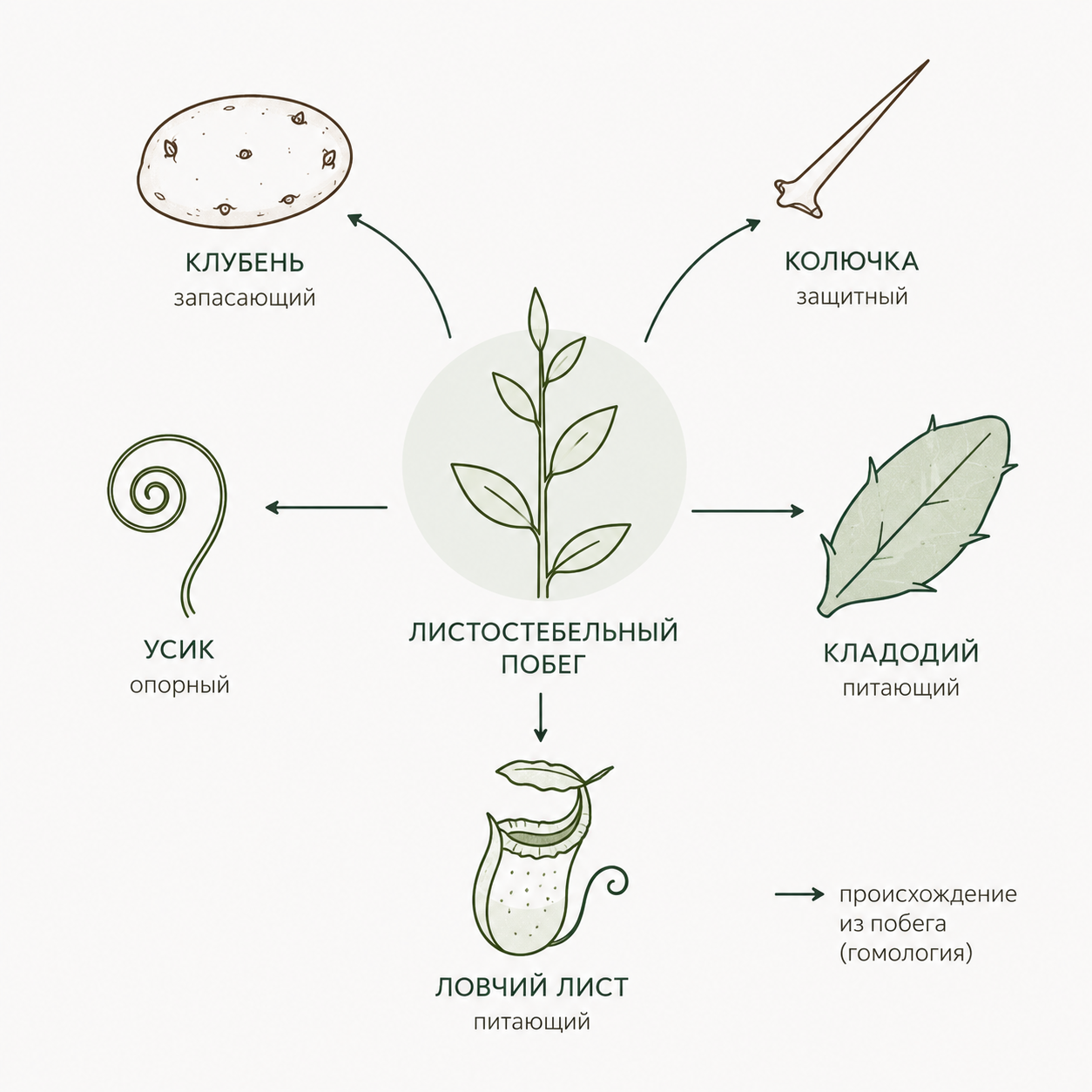 Рис. 1. Схема развития листостебельного побега.
Рис. 1. Схема развития листостебельного побега.
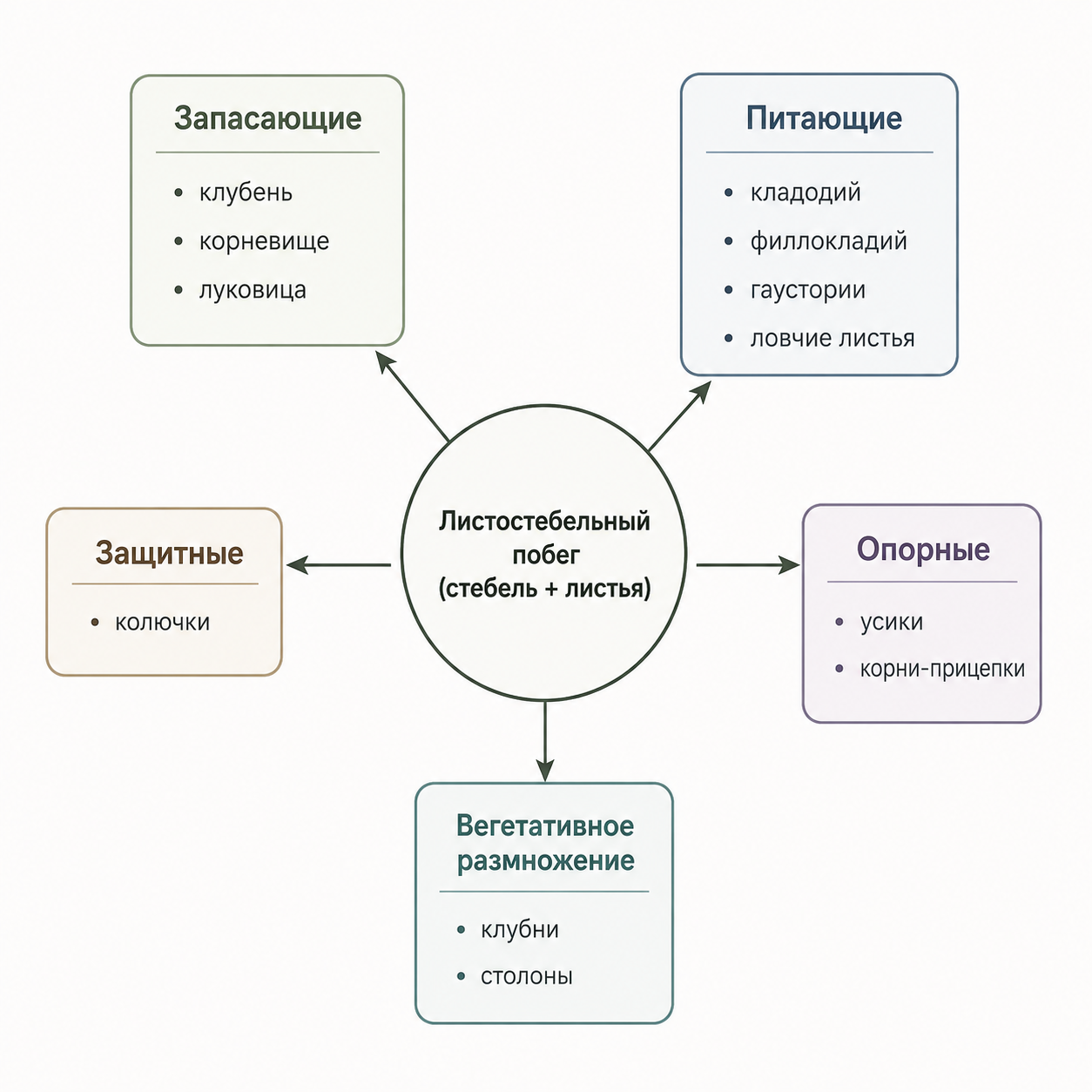 Рис. 2. Варианты развития побега.
Рис. 2. Варианты развития побега.
Эволюция и анатомия стебля
Ранний этап формирования стебля начинается с семядоли, под которой располагается гипокотиль, находящийся в непосредственном контакте с зародышевым корнем. На поверхности многолетнего корневища ясно видны рубцы от прошлогодних низовых листьев и отмерших побегов, а на самом начальном этапе развития присутствует отдельный рубец от семядоли. Эти остаточные следы позволяют проследить последовательную морфогенетическую трансформацию от зародышевой стадии до зрелого вегетативного органа.
У споровых растений — плаунов, папоротников и хвощей — стебли характеризуются особым конусом нарастания, в центре которого находится стела — центральная часть осевого цилиндра. Такая организация представляет собой раннюю форму стеблевой системы, от которой эволюционировали более сложные конструкции у семенных растений. У голосеменных и двудольных покрытосеменных конус нарастания усложняется: уже на ранних стадиях закладывается прокамбий, который дифференцируется в первичные проводящие ткани. Общий план эвстелы, типичный для этих групп, обеспечивает эффективный транспорт материалов и служит важной эволюционной особенностью.
Эвстела проявляет разнообразие в организации открытых проводящих пучков, расположенных вокруг сердцевины, либо в виде трубчатого цилиндра ксилемы, окружённого цилиндром флоэмы. В зависимости от типа ствола она может быть пучковой (с широкими лучами сердцевины) или непучковой (со сплошной ксилемой и узкими лучами). Отличие эвстелы от сифноствелы состоит в способности к вторичному утолщению за счёт развития камбия.
Эпидерма стеблей двудольных растений сохраняет обычное строение с хорошо развитыми устьицами и разнообразными волосками, а первичная кора и сердцевина в комбинации с вариациями эвстелы формируют широкий спектр анатомических особенностей. В узлах, где прикрепляются листья и закладываются почки, наблюдается специализированное строение, отличающееся от междоузлий; эти зоны служат центрами ветвления и механической прочности. Стебли однодольных растений отличаются от двудольных отсутствием настоящей камбиальной зоны, что лишает их возможности вторичного утолщения. Проводящие пучки у однодольных рассредоточены по всему стеблю, что является характерным диагностическим признаком.
Вторичное утолщение у палеозойских и современных древесных растений происходит различными механизмами, отражая эволюционные изменения в адаптации к наземному образу жизни. Несмотря на различия в долговечности, однолетние стебли, многолетние ветви и стволы хвойных и лиственных деревьев демонстрируют общие принципы: камбий, как слой делящихся клеток, образует наружные ткани (флоэму) и внутренние (ксилему), формируя годичные кольца, служащие важными таксономическими маркерами.
Метаморфозы стебля и проводящих систем
В процессе вторичного утолщения клетки прокамбия одновременно закладывают будущие элементы вторичной флоэмы и ксилемы, а также продолжают дифференцировать первичные проводящие ткани. После завершения этой первичной дифференциации дальнейшее увеличение объёма проводящих систем осуществляется исключительно камбием, образующим годичные слои. При формировании коллатерального проводящего пучка первичная флоэма образуется из наружных клеток прокамбия, а первичная ксилема закладывается эндочно. Такой открытый пучок отличается от закрытого, содержащего лишь первичные ткани; вторичные ткани развиваются между ними, отодвигая флоэму к периферии. Биколлатеральный пучок формируется аналогично, но с дифференциацией флоэмы и из внутренних, и из наружных клеток прокамбия, образуя два кольца — флоэму и ксилему — вложенных друг в друга.
Метаморфозы стебля проявляются в таких формах, как кочан обычной столовой капусты, где огромная почка образует мясистый стебель-кочерыгу, поддерживая сочные листовые пластины. У брюссельской капусты формируются мелкие кочаны из боковых почек, что иллюстрирует вариативность преобразований. Ещё один пример — корневище, представляющее собой подземный побег с чешуевидными листьями, почками и придаточными корнями. На его поверхности видны рубцы от прошлогодних листьев и отмерших побегов, что свидетельствует о его роли в выживании и вегетативном размножении в неблагоприятных условиях.
Эволюция ветвления начинается с равной дихотомии (изотомии), когда обе ветви растут синхронно. Затем она переходит к неравной дихотомии (анизотомии), при которой одна ветвь опережает другую, что может привести к дихоподиальному ветвлению с зигзагообразным ростом главной оси. Из дихотомического ветвления развились моноподиальное и симподиальное типы боковых ветвлений. При моноподиальном росте главная ось сохраняет неограниченный верхушечный рост, а боковые ветви уступают ей. При симподиальном росте главная ось прекращает удлинение, и ближайшая боковая ветвь заменяет её, смещая конец роста.
Теломы — цилиндрические органы косо-вертикальной ориентации — предшествовали современным стеблям. Их увеличение в размерах и последующее уплощение в листовые структуры, ориентированные горизонтально, позволило расширить светоприёмную поверхность и стало ключевым шагом в эволюции вегетативных органов высших растений. По дарвиновской эволюционной теории, метаморфоз в растениях представляет собой не случайную морфологическую модификацию, а генетически закреплённый процесс трансформации органов, обусловленный изменяющимися условиями среды и тесно связанный с историческим развитием вида и его адаптацией к новым экологическим нишам.
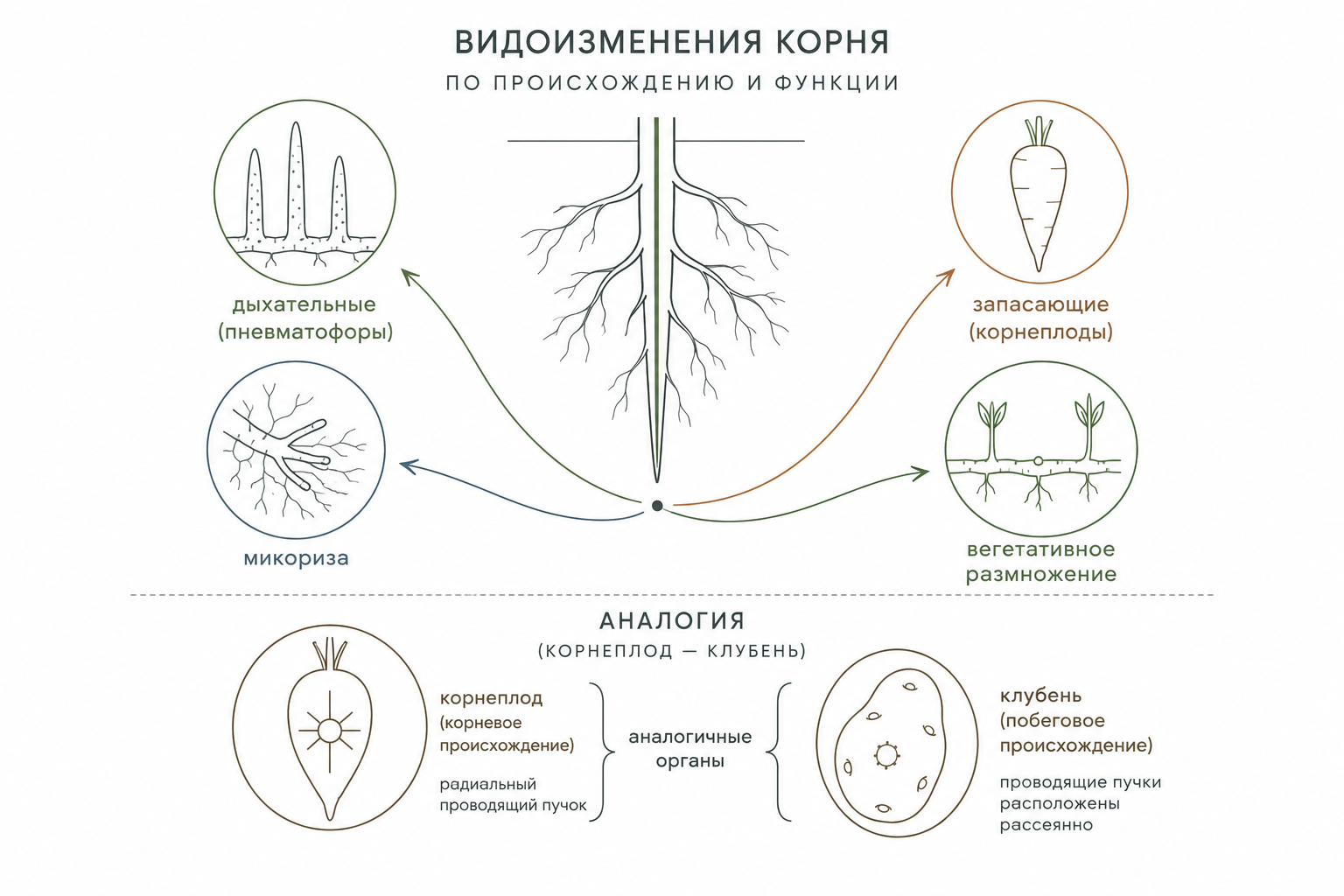
Видоизменения корневой системы
Дыхательные корни, или пневматофоры, представляют собой специализированные ветвления корневой системы, отличающиеся отрицательным геотропизмом и ростом вертикально вверх. У субтропических и тропических растений они образуются из длинных горизонтальных подземных корней. У болотного кипариса такие пневматофоры выступают над поверхностью почвы в виде пеньков высотой до нескольких десятков сантиметров. Внешне их покрывает перидерма с характерными чечевичками, а внутри развитая губчатая ткань, межклетники которой заполнены воздухом, обеспечивает эффективный газообмен. У водного южноамериканского растения жюссеи ползучей верхушки беловатых пневматофоров также выступают над водой, где служат основным механизмом дыхания. В их корне центральный цилиндр развит слабо, но первичная кора мощна и состоит из аэренхимы с обильными воздухоносными полостями.
Метаморфозы корней классифицируются как по их происхождению из разных вегетативных органов, так и по функциональному принципу. Функциональные метаморфозы включают запасающие формы, в которых органические вещества, вода и воздух сосредоточены в межклетниках и крупных полостях. Функциональная специализация корневых метаморфозов проявляется в соотношении сосудистых тканей: в запасающих формах флоэма обычно менее развита, тогда как ксилема лучше дифференцирована, что обеспечивает транспорт синтезированных веществ к видоизменённому органу.
Органы, сходные по функции и морфологии, но различающиеся по происхождению, называют аналогичными. Например, корневые шишки и побеговые клубни оба представляют собой мясистые структуры с запасом питательных веществ, однако в первых сохраняется радиальное расположение экзархной первичной ксилемы, а во вторых — типичная стеблевая проводящая система. Аналогия часто сопровождается редукцией: развитие колючек барбариса сопровождается редукцией мезофилла. Именно такие аналогичные конструкции становятся основой конвергенции — независимого появления сходных морфологических признаков у растений разных систематических групп. Ярким примером служит кактусообразный облик пустынных растений разных семейств (кактусовые, молочайные, сложноцветные, ластовневые), у которых сочные стебли изобилуют слизистыми паренхимными клетками, удерживающими воду.
Корневая природа любого метаморфоза всегда определяется наличием центрального цилиндра с радиальным проводящим пучком и экзархной первичной ксилемой, независимо от внешних модификаций. У люцерны посевной главный корень может достигать 10–15 м, позволяя глубоко проникать в почву. У большинства однодольных и некоторых двудольных растений главный корень быстро отмирает, а система состоит преимущественно из многочисленных придаточных корней, образующих мочковатую структуру. В древесных видах корни функционально дифференцированы: долговечные скелетные корни расширяют площадь питания и закрепляют растение, тогда как короткие сосущие корни отвечают за поглощение воды и минералов. Сосущие корни вступают в микоризу — симбиотическое взаимодействие с грибами. Она может быть экзотрофной (гифы оплетают корень снаружи) или эндотрофной (проникают внутрь и образуют клубок). Такие корни не имеют корневых волосков, их апикальный рост прекращается рано, а особое ветвление максимизирует площадь контакта с грибными гифами.
Интенсивность развития корневой системы определяется условиями среды: у мятлика однолетнего на тяжёлых глинистых почвах корни достигают лишь 10 см, а на прибрежных песках — 25–30 см. У многих древесных растений (ели, сосны, дуба, берёзы) и некоторых многолетних трав корневая система представляет собой смешанный тип. Среди водных растений существуют специальные корни, адаптированные к водной среде для дыхания и фиксации. Корни также видоизменяются в органы запасания (корнеплоды у моркови, свёклы, картофеля) или вегетативного размножения (пырей, бамбук).
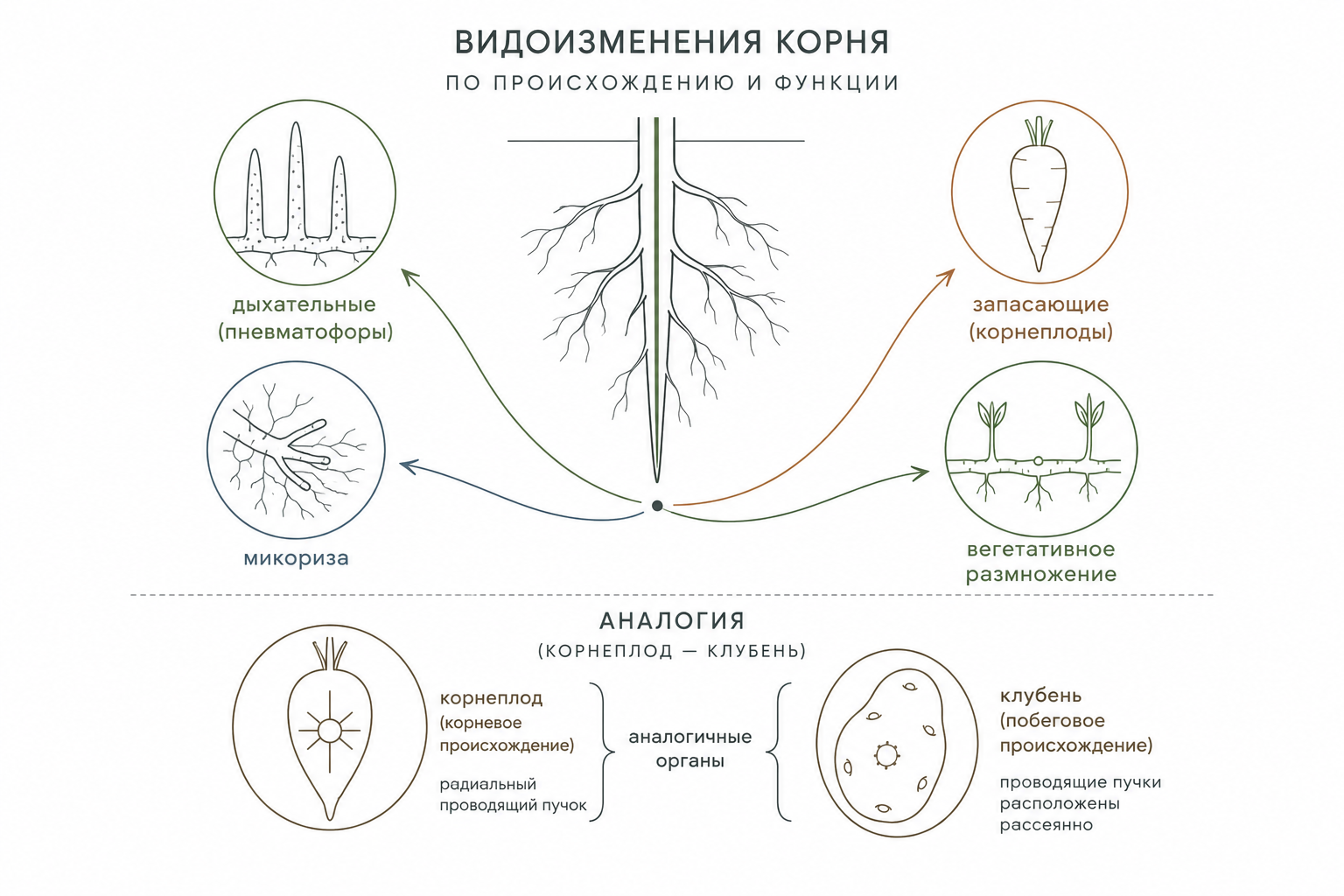 Рис. 3. Схема классификации метаморфозов корней.
Рис. 3. Схема классификации метаморфозов корней.
Строение и метаморфозы листа
Пластинка листа — центральный орган растения, в котором совершаются фотосинтез, транспирация и дыхание. Текстурные особенности пластинок отражают адаптивные стратегии: у большинства растений они тонкие и мягкие, у падуба и самшита — кожистые, у эфедры и казуарины — пленчатые, а у агавы, молодила и эхеверии — сочные и мясистые. Зелёный цвет обусловлен присутствием хлорофилла, однако его спектр иногда маскируется другими пигментами, например антоцианами. Серебристо-белые пятна появляются там, где клетки заменяют хлоропласты бесцветными лейкопластами или содержат множество межклетников, заполненных воздухом, который рассеивает лучи.
В сложных листах пластинок несколько, и каждая обычно имеет собственный черешок, отходящий от общего черешка-рахиса. Различают тройчато-сложные, пальчато-сложные (веером) и перистосложные (в два ряда вдоль рахиса) листья. Гетерофиллия проявляется, когда у одной особи форма и размеры листьев различаются: у водяного лютика надводные и подводные листья имеют разное строение. У папоротников рода Platycerium сначала формируются крупные цельные листья, прижатые к стволу дерева; между стволом и листом образуется ниша, где накапливается гумус, в который впадают придаточные корни. Листовые зачатки закладываются на конусе нарастания экзогенно, формируясь из фланговой зоны и мантии (туники).
Метаморфоз листовых органов принимает широкого спектра форм. Почечные чешуи, являющиеся низовыми листьями, постепенно переходят в полноценные листовые пластины по мере роста побега. Верховые листья, покрывающие цветки и соцветия, могут быть мелкими и пленчатыми или крупными и ярко окрашенными (как у белокрыльника, антуриумов, бромелиевых и марьянника дубравного), выполняя функцию привлечения опылителей. У морозника выражена ярусная дифференциация: от низовых семядолей через срединные ассимилирующие до переходных и верховых листьев с редуцированными пластинками. Почки, будучи точками роста, формируют все вегетативные органы из конуса нарастания главного побега. Почечные чешуи защищают зарождающиеся органы, представляя собой редуцированные листья, сменившие ассимиляционную функцию на защитную.
Побег в целом рассматривается как целостная структурно-функциональная система, способная к множественной лабильности. Кочан обычной столовой капусты можно считать гигантской почкой-запасателем, где мясистый стебель (кочерыга) образует центральную часть. Аналогично, корневище представляет собой побег с чешуевидными листами, почками и придаточными корнями. На поверхности многолетнего корневища видны рубцы от прошлогодних листьев и отмерших побегов, а в ранней стадии развития наблюдается рубец от семядоли, под которой находится гипокотиль, граничащий с зародышевым корнем.
Листопад играет важнейшую экологическую роль, позволяя растениям преодолевать неблагоприятные периоды и предотвращая иссушение зимой. При низких температурах корневая система слабо поглощает воду, поэтому опад листьев стал адаптивным механизмом в ответ на чередующиеся климатические условия.
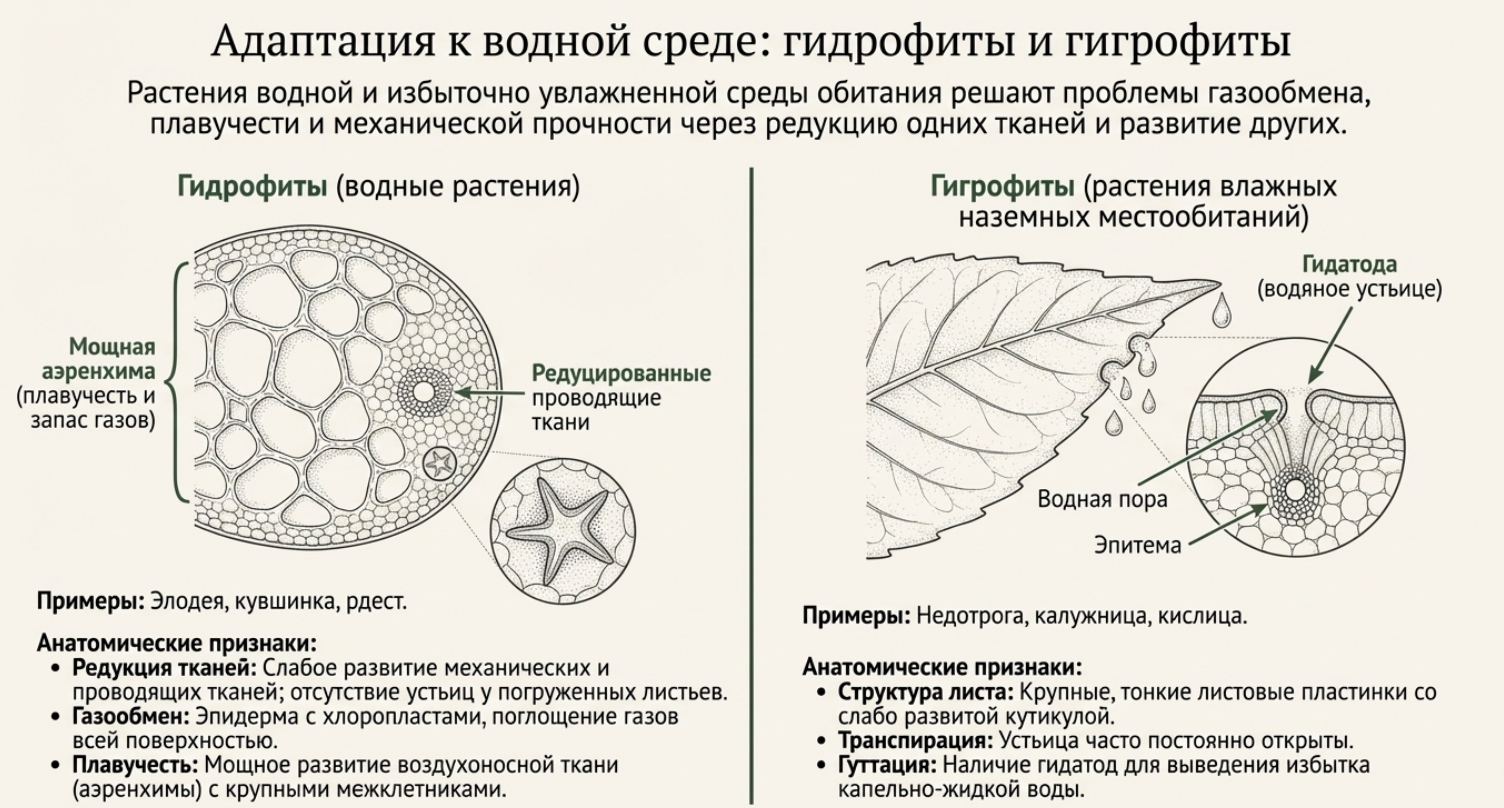
Экологические адаптации: ксерофиты и гидрофиты
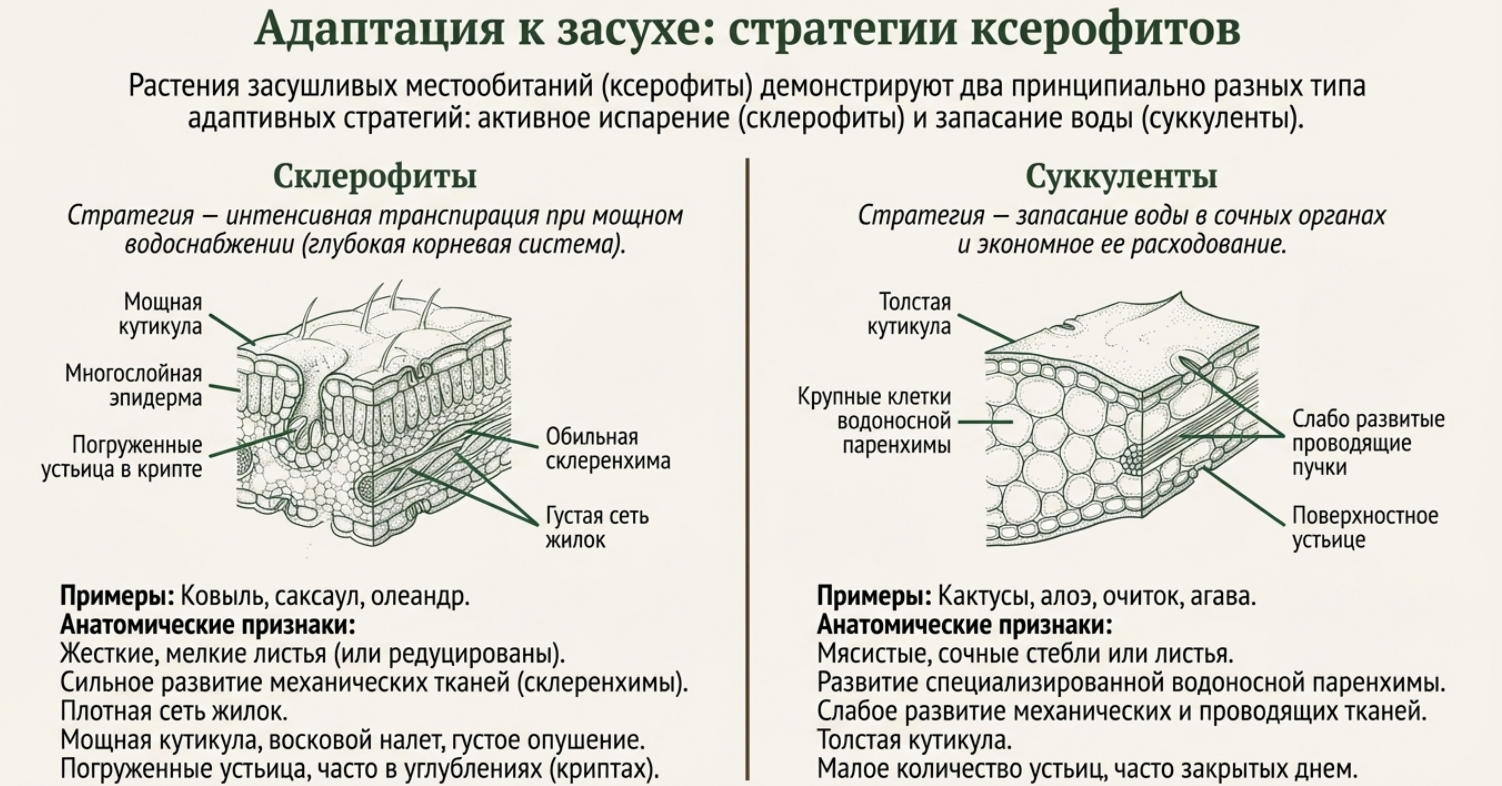
Ксерофиты, обитающие в экстремально сухих местообитаниях, поддерживают жизнедеятельность при дефиците влаги благодаря комплексу морфологических и анатомических приспособлений. Их корневая система может достигать глубины тридцати метров и более (верблюжья колючка, саксаул). Стебли и листья ксерофитов эластичны и сильно редуцированы, что снижает испарительные потери, а желёзистые ткани удерживают воду внутри паренхимных клеток. Кератинизация и усиленная кальцификация зубчатых колючек кактусов представляют собой метаморфоз почечных листьев в органы, одновременно снижающие транспирацию, конденсирующие водяные пары и защищающие от травоядных. Суккулентные стебли наполнены слизью, позволяя растениям функционировать без листьев или при их преждевременном опадении.
Гидрофиты, в отличие от наземных ксерофитов, погружены в водный столб и, как правило, не развивают мощной корневой системы. Их корневые волоски часто отсутствуют, а поглощение воды и минералов происходит через поверхность листьев и стеблей. В эпидерме плавающих листьев располагаются специальные клетки — гидропоты, активно всасывающие и выделяющие воду. Листья покрыты верхними слоями бесцветных паренхимных клеток, содержащих жироподобные вещества, которые отражают избыточное тепловое излучение. Подводные листья часто обладают удлинённой тесьмовидной формой (валлиснерия) или сильно разрезаны на тонкие сегменты (уруть, роголистник, пузырчатка, водные лютики), что увеличивает фотосинтезирующую площадь. У водяного ореха наблюдается метаморфоз корней в сильно разрезанные ассимилирующие органы. Железистые волоски и листовые кристаллы служат адаптивными структурами для выживания в экстремальных условиях у обеих экологических групп.
Подробнее об адаптациях к засушливой и влажной среде
Заключение
Метаморфозы классифицируются по двум основным признакам: по происхождению из того или иного вегетативного органа и по функциональному принципу. Эта систематизация позволяет упорядочить огромное разнообразие видоизменений растений. По дарвиновской эволюционной теории, метаморфоз — это не случайная модификация, а генетически закреплённый процесс трансформации, обусловленный изменяющимися условиями среды и тесно связанный с историческим развитием вида. Побег представляет собой целостную структурно-функциональную систему, способную к множественной лабильности. Главные и боковые побеги, почки и листья способны метаморфизироваться в ответ на экологические вызовы. Независимо от того, формируется ли гигантская почка-запасатель, корневище с редуцированными чешуевидными листьями или дыхательные пневматофоры, растения демонстрируют безграничные возможности адаптивной эволюции, оптимизируя свою форму и функцию ради выживания в постоянно меняющемся мире.