Надпочечники и их роль в эндокринной системе
Надпочечники — парные эндокринные железы, расположенные над верхними полюсами почек и покрытые плотной соединительнотканной капсулой. У взрослого человека их масса составляет около 4–6 г. Несмотря на скромные размеры, эти железы выполняют колоссальную работу: они синтезируют десятки гормонов, управляющих обменом веществ, водно-солевым равновесием, адаптацией к стрессу и формированием половых признаков. Под капсулой находится корковый слой, занимающий примерно 15 % площади железы, тогда как оставшаяся часть приходится на более массивное мозговое вещество, формирующее внутреннюю часть надпочечника. Такое архитектурное деление неслучайно: каждая зона вырабатывает собственный набор стероидов или катехоламинов, действуя как самостоятельная, но тесно интегрированная эндокринная фабрика.
Строение и организация коры надпочечников
Кора надпочечников состоит из трёх морфофункциональных зон, расположенных последовательно от периферии к центру: наружной клубочковой (зона сафра), средней пучковой (зона фасцикулярная) и внутренней сетчатой (зона ретикулярная). Каждая зона обладает характерным набором специализированных клеток и синтезирует определённые группы стероидных гормонов, что обеспечивает чёткое функциональное разделение труда внутри одного органа.
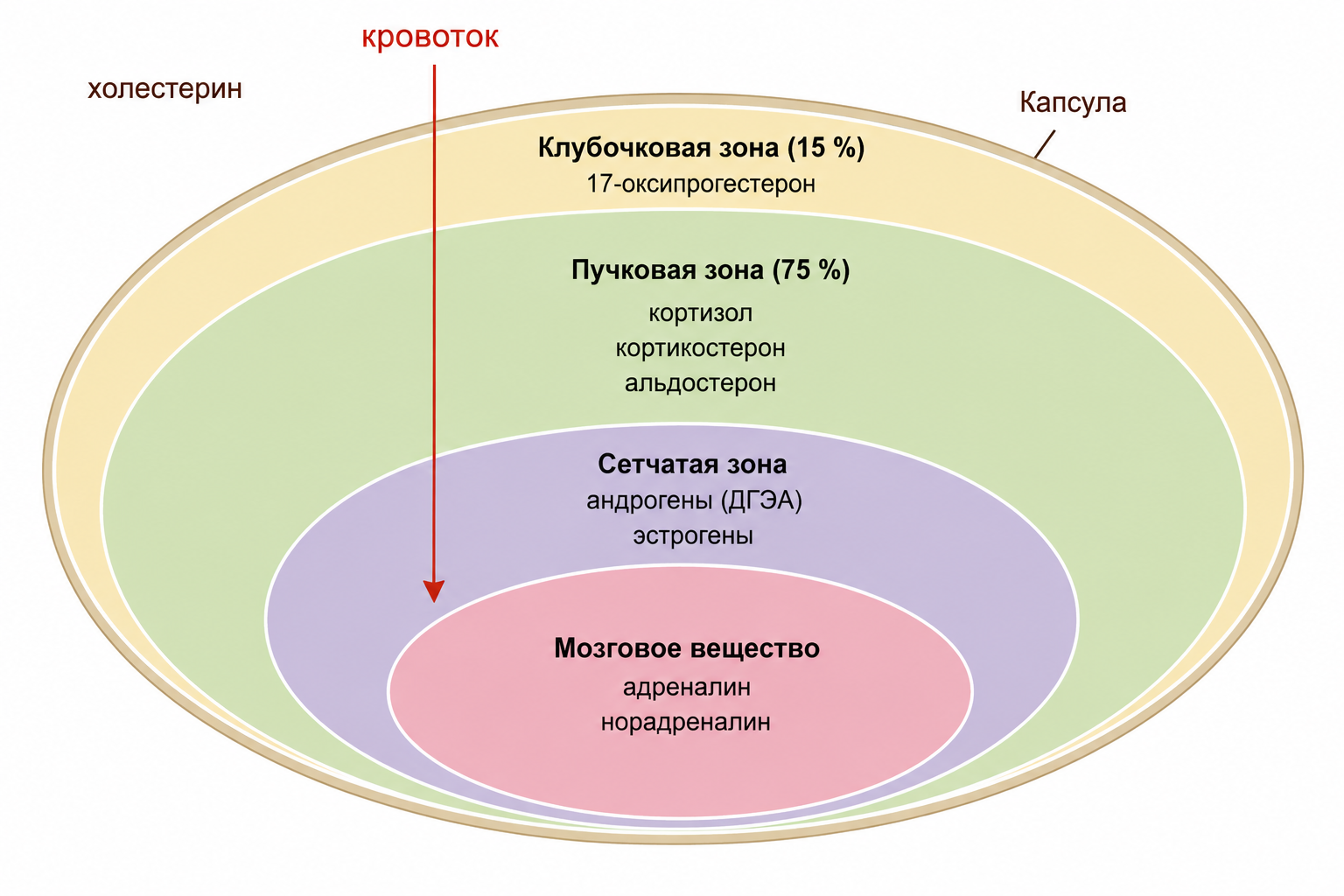 Рис.1. Схема поперечного среза надпочечника.
Рис.1. Схема поперечного среза надпочечника.
Клубочковая зона, являясь самой наружной, составляет около 15 % толщины коры. Клетки этой зоны способны синтезировать только предшественники стероидов, такие как 17-оксипрогестерон, но не завершают путь до активных гормонов, поскольку в их ферментном наборе отсутствуют необходимые ферменты для дальнейшего преобразования. Это означает, что зона работает как поставщик промежуточных метаболитов, окончательная «сборка» которых происходит в других отделах коры.
Пучковая зона, занимающая примерно 75 % толщины коры, является самой широкой. Её клетки продуцируют глюкокортикоиды (кортизол, кортикостерон) и минералокортикоиды (альдостерон). Здесь высока плотность митохондрий, а в их мембранах находятся ферменты (например, 11β-гидроксистероиддегидрогеназа), отвечающие за окончательную стадию синтеза кортизола. Благодаря такому обилию митохондрий пучковая зона способна мгновенно реагировать на сигналы АКТГ мощным выбросом глюкокортикоидов.
Сетчатая зона, расположенная непосредственно под пучковой, отвечает за выработку андрогенов (дегидроэпиандростерон и др.) и небольших количеств эстрогенов. Клетки этой зоны характеризуются повышенным содержанием липидных капель, где хранится предшествующий стероидный субстрат. Таким образом, даже небольшая по объёму зона способна длительно и стабильно поставлять половые гормоны надпочечникового происхождения.
Мозговое вещество надпочечника (медулла) находится в центральной части железы и состоит из хромаффинных клеток, которые быстро вырабатывают катехоламины — адреналин и норадреналин. Их секреция происходит в ответ на симпатическую активацию и обеспечивает быструю реакцию «борьбы-бегства». В отличие от коры, действующей минутами и часами, мозговое вещество отвечает на стресс за секунды.
Гистологически клетки коры надпочечника отличаются высокой плотностью митохондрий, наличием липидных капель (более выраженным в зонах, синтезирующих стероиды) и специфической границей с капиллярной сетью, через которую доставляется холестерин — предшественник всех стероидных гормонов. Слой капилляров, расположенный сразу под капсулой, образует сеть, в которой кровь поступает к каждой зоне коры через уникальные сосудистые артерии (корковые артерии) и выходит из зоны через венулы, обеспечивая градиентный обмен субстратов и гормонов между зонами.
Функциональная связь между корой и мозговым веществом обеспечивается внутрижелезным нервным волокном, которое передаёт симпатическую импульсную информацию от спинного мозга к хромаффинным клеткам, вызывая мгновенный всплеск катехоламинов при стрессовых ситуациях. Эта нейроэндокринная связь делает надпочечник уникальным органом, в котором гормональная и нервная регуляция физически сопряжены.
Интересно, что у разных видов реакция надпочечников на острый стресс может существенно различаться. У крыс, например, реакция на острый стресс (перелом конечностей) проявляется в ускоренной секреции кортикостерона вместо кортизола, что свидетельствует о гибкости зонирования гормональных путей в ответ на физиологический нагрузочный стимул. Подобные межвидовые различия подчёркивают, что набор ферментов в коре надпочечников эволюционно пластичен и подстраивается под условия среды обитания.
Более подробная схема надпочечников
 Antinksčio sandara.png Author: EdgarasLe, CC BY-SA 4.0 https://creativecommons.org/licenses/by-sa/4.0, via Wikimedia Commons
Antinksčio sandara.png Author: EdgarasLe, CC BY-SA 4.0 https://creativecommons.org/licenses/by-sa/4.0, via Wikimedia Commons
Стероидогенез: путь от холестерина к гормонам
Холестерол (строительный материал для гормонов) попадает в клетки коры надпочечников из крови в составе ЛПНП. Гормон АКТГ заставляет клетки активнее захватывать эти частицы и извлекать из них холестерол. Это одновременно и «закупка» сырья, и сигнал к его переработке.
Первый шаг — превращение холестерола в вещество прегненолон. Это самая медленная стадия, именно она задаёт темп всему процессу. В пучковой зоне коры (где делается кортизол) её ускоряет АКТГ, а в клубочковой зоне (где делается альдостерон) — ангиотензин II.
Дальше прегненолон превращается в разные гормоны в зависимости от зоны:
В пучковой зоне — цепочка реакций ведёт к кортизолу.
В клубочковой зоне — уникальный фермент, работающий только здесь, направляет процесс к альдостерону.
В сетчатой зоне — из того же сырья получается дегидроэпиандростерон (ДГЭА) — главный половой гормон надпочечников.
Если какого-то фермента в этих цепочках не хватает, гормональный баланс сильно нарушается. Например, при недостатке фермента на пути к кортизолу сырьё уходит в обходной путь, вызывая избыток половых гормонов (адреногенитальный синдром).
Количественные факты:
Кортизол даёт ≈95% всей глюкокортикоидной активности, его концентрация в крови — 12 нг/дл, выделяется 15 мг в сутки.
Кортикостерон выделяется 3 мг/сут, но он втрое слабее кортизола.
Альдостерон, несмотря на ничтожную концентрацию (0,006 нг/дл) и секрецию всего 0,15 мг/сут, обеспечивает ≈90% минералокортикоидной активности. Причина: его сродство к рецепторам в 3000 раз выше, чем у кортизола. Сам кортизол в почках быстро разрушается специальным ферментом, поэтому почти не действует как минералокортикоид.
Дезоксикортикостерон секретируется примерно столько же, сколько альдостерон, но его активность — лишь 1/30 активности альдостерона.
Синтетические аналоги мощнее природных:
Дексаметазон — в 30 раз активнее кортизола как глюкокортикоид.
9α-флюорокортизол — в 125 раз активнее альдостерона как минералокортикоид.
Преднизолон и метилпреднизон — соответственно в 4 и 5 раз активнее кортизола.
Такие различия объясняются тонкими особенностями строения молекул: именно они определяют, с какими рецепторами гормон свяжется и насколько сильно подействует.
Более подробно о стероидогенезе
Транспорт холестерола в надпочечники происходит через рецептор-опосредованный эндоцитоз липопротеинов низкой плотности (ЛПНП). ЛПНП, обогащённые холестеролом, покидают плазму крови, проникают в интерстициальное пространство и фиксируются на рецепторах, расположенных в окаймлённых ямках мембран адренокортикальных клеток. После связывания ямки погружаются внутрь клетки, образуя везикулы, которые в конечном итоге сливаются с лизосомами; в результате холестерол высвобождается и становится доступным для стероидогенеза. Количество доступного холестерола регулируется в значительной мере обратной связью: адренокортикотропный гормон (АКТГ) повышает экспрессию рецепторов ЛПНП на поверхности клеток и одновременно активирует ферменты, высвобождающие холестерол из ЛПНП, тем самым усиливая как захват, так и мобилизацию холестерола. Таким образом, клетка одновременно «закупает» сырьё и запускает его переработку.
Лимитирующей стадией синтеза стероидов является превращение холестерола в прегненолон под действием холестеролдесмолазы (цитохром P450scc) в митохондриях. Попавший в клетку холестерол быстро транспортируется в митохондрии, где фермент расщепляет его с образованием прегненолона; скорость этого шага определяет общий темп стероидогенеза. В разных зонах коры надпочечников лимитирующая стадия стимулируется различными регуляторами: в зонах, отвечающих за глюкокортикоиды, её ускоряет АКТГ, а в клубочковой зоне коры, где синтезируется альдостерон, активность повышает ангиотензин II. Следовательно, один и тот же биохимический узел становится точкой интеграции различных физиологических сигналов, обеспечивая дифференцированную продукцию глюкокортикоидов и минералокортикоидов.
После образования прегненолона дальнейшие модификации происходят в эндоплазматическом ретикулуме, где задействованы ферменты суперсемейства цитохромов P450. Путь к кортизолу включает последовательные реакции: прегненолон → 17α-гидроксипрегненолон (катализируется 17α-гидроксилазой, P450c17) → 17α-гидроксипрогестерон → 11-дезоксикортизол (P450c21) → кортизол (P450c11). Наличие 17α-гидроксилазы определяет способность пучковой зоны коры синтезировать глюкокортикоиды; её дефицит приводит к блокировке пути. Синтез альдостерона отклоняется от общего пути уже на уровне прогестерона и проходит через стадии прогестерон → 11-дезоксикортикостерон → кортикостерон → альдостерон, причём ключевую роль играет альдостеронсинтаза (P450c11AS), экспрессирующаяся исключительно в клубочковой зоне коры, что объясняет локализованный характер минералокортикоидного синтеза. В сетчатой зоне коры активность 17α-гидроксилазы наиболее высока, что позволяет 17α-гидроксипрегненолону через 17,20-лиазную активность того же фермента P450c17 образовывать дегидроэпиандростерон — основной андроген надпочечникового происхождения.
Изменения активности любого из ферментов цепи приводят к существенным колебаниям как количества, так и относительной доли отдельных гормонов. Повышение активности 21β-гидроксилазы, например, может вызвать избыточную продукцию половых гормонов и маскулинизацию, тогда как её дефицит приводит к клинически значимому синдрому с нарушением минерального и глюкокортикоидного баланса. Чувствительность системы такова, что даже небольшое колебание активности одного фермента меняет весь гормональный профиль пациента.
Количественные параметры секреции подчёркивают разницу между гормонами. Кортизол, обеспечивая около 95 % глюкокортикоидной активности, имеет среднюю плазменную концентрацию 12 нг/дл и скорость секреции 15 мг/сут. Кортикостерон, хотя и секретируется лишь в количестве 3 мг/сут, обладает глюкокортикоидной активностью 0,3 от активности кортизола. Альдостерон, несмотря на плазменную концентрацию всего 0,006 нг/дл и скорость секреции 0,15 мг/сут, обеспечивает почти 90 % минералокортикоидной активности благодаря своему 3000-кратному аффинитету к минералокортикоидным рецепторам. Кортизол, хотя и присутствует в плазме в тысячи раз выше, обладает лишь единичной минералокортикоидной активностью, поскольку в мишенях он быстро инактивируется 11β-гидроксистероиддегидрогеназой 2. Дезоксикортикостерон, с концентрацией и уровнем секреции, сопоставимыми с альдостероном, вносит лишь незначительный вклад (1/30 активности альдостерона).
Синтетические стероиды демонстрируют гораздо более выраженные эффекты: дексаметазон обладает глюкокортикоидной активностью в 30 раз выше кортизола, 9α-флюорокортизол — минералокортикоидной в 125 раз, преднизолон — в 4 раза, а метилпреднизон — в 5 раз. Их действие основано на химических особенностях молекул: у кортизола кетогруппа в C3 и гидроксильные группы в C11 и C21, тогда как у альдостерона C18 образует альдегидную группу, участвующую в полуацетальном цикле. Эти различия определяют избирательность к глюкокортикоидным и минералокортикоидным рецепторам и объясняют, почему даже небольшие структурные модификации радикально меняют профиль активности гормона.
Кортизол — основной глюкокортикоид организма
Кортизол (гидрокортизон) является основным глюкокортикоидом в организме человека, обеспечивая около 95 % всей глюкокортикоидной активности от общего уровня продукции глюкокортикоидов. Дополнительный, но функционально значимый вклад вносит кортикостерон, который обладает слабой минералокортикоидной активностью и секретируется в очень малом количестве. Глюкокортикоиды столь же важны для нормальной продолжительности жизни, как и минералокортикоиды: животные, лишённые этих гормонов, не способны противостоять различным видам физических и даже умственных стрессов, и такое, казалось бы, рядовое событие, как респираторная инфекция, может привести к гибели при недостатке глюкокортикоидов.
Один из главных эффектов кортизола — стимуляция глюконеогенеза в печени, то есть процесса образования углеводов из белков и некоторых других веществ; кортизол способен повышать уровень глюконеогенеза в 6–10 раз. Этот эффект обеспечивается двумя основными механизмами: увеличением количества ферментов для превращения аминокислот в глюкозу и мобилизацией аминокислот из внепечёночных тканей. Кортизол увеличивает количество ферментов, необходимых для превращения аминокислот в глюкозу в клетках печени, что является результатом активации глюкокортикоидами процессов транскрипции ДНК — подобно тому как альдостерон активирует их в тубулярных клетках почек, — с последующим образованием мРНК, ведущим к увеличению спектра ферментов, необходимых для глюконеогенеза.
Параллельно кортизол способен мобилизовать аминокислоты из внепечёночных тканей, главным образом из мышц. В результате возрастает количество аминокислот, которые могут поступать в печень и служить сырьём для образования глюкозы. Одним из проявлений возросшего глюконеогенеза под действием кортизола является увеличение запасов гликогена в клетках печени. Это влияние кортизола позволяет гликолитическим гормонам, таким как адреналин, мобилизовывать глюкозу по мере необходимости, например в промежутках между приёмами пищи. Кроме того, кортизол умеренно снижает уровень потребления глюкозы клетками, что способствует повышению концентрации глюкозы в крови и обеспечению энергетических потребностей организма в условиях стресса.
Кортизол способствует умеренно выраженной мобилизации жирных кислот из жировой ткани, что увеличивает их концентрацию в плазме крови. Механизм мобилизации жирных кислот кортизолом не вполне понятен, однако часть эффектов опосредована снижением поступления глюкозы в жировые клетки. Поскольку α-глицерофосфат (один из дериватов глюкозы) необходим для поступления и сохранения триглицеридов в клетках, его отсутствие в адипоцитах ведёт к высвобождению жирных кислот. Увеличение мобилизации жиров кортизолом наряду с увеличением окисления жирных кислот в клетках обеспечивает сдвиг метаболических систем в сторону использования жирных кислот вместо глюкозы в качестве источника энергии при голодании и других стрессах. Этот механизм требует для своего формирования несколько часов и не является столь же быстрым и мощным, как эффект похожего сдвига, вызванного снижением инсулина.
Избыток кортизола вызывает своеобразный тип ожирения с избыточным отложением жира в шейно-воротниковой зоне, создающим впечатление «бычьего торса», и на лице (так называемое «лунолицо»). Хотя причины такого перераспределения жира неизвестны, предполагается, что этот тип ожирения является результатом мощной стимуляции потребления пищи, когда жиры в некоторых тканях образуются быстрее, чем мобилизуются или окисляются. Физический или нейрогенный стресс вызывает незамедлительное и заметное увеличение продукции АКТГ передней долей гипофиза в течение нескольких минут, приводящее к резкому увеличению секреции кортизола корой надпочечников. Таким образом, глюкокортикоиды играют ключевую роль в адаптации организма к стрессовым воздействиям.
Противовоспалительное и противоаллергическое действие кортизола
Кортизол обладает мощным противовоспалительным действием, способным уменьшить воспаление в течение нескольких часов или дней. Быстрый эффект связан с блокадой большинства факторов, формирующих воспаление; кроме того, повышается скорость заживления. Противовоспалительное действие кортизола может быть результатом нескольких механизмов: мобилизации аминокислот и их использования для восстановления повреждённых тканей; возрастания уровня глюконеогенеза, обеспечивающего систему избытком глюкозы в критических условиях метаболизма; увеличения количества жирных кислот, необходимых для энергообеспечения клеток; а также эффекта кортизола, инактивирующего и устраняющего продукты воспаления.
Кортизол играет главную роль в борьбе с такими заболеваниями, как ревматоидный артрит, ревматическая атака или острый гломерулонефрит. Данные болезни характеризуются тяжёлым местным воспалением, и их повреждающее действие обусловлено в основном воспалением как таковым. Если назначить кортизол или другие глюкокортикоиды больным с воспалительными заболеваниями, проявления воспаления пойдут на убыль в течение суток. Даже если кортизол не действует на главную причину заболевания, а только предупреждает повреждающий эффект воспалительного ответа, один этот фактор может спасти жизнь человека.
Помимо собственно воспаления, кортизол блокирует воспалительные компоненты аллергических реакций. Он не блокирует взаимодействие антигена и антитела, лежащее в основе аллергической реакции, и вторичные проявления аллергии сохраняются, однако воспалительные компоненты, ответственные за многие серьёзные, а иногда и летальные исходы аллергических реакций, ослабляются кортизолом. Введение кортизола и опосредованное им уменьшение проявлений воспаления наряду со снижением высвобождения продуктов воспаления может быть спасительным при аллергических реакциях. Так, кортизол во многих случаях предупреждает развитие шока и смерть при анафилаксии.
Транспорт кортизола и синтетические глюкокортикоиды
Приблизительно 90–95 % кортизола в плазме связаны с белками, особенно с глобулином, названным кортизол-связывающим глобулином, или транскортином, и в меньшей степени — с альбумином. Высокая белоксвязывающая способность кортизола уменьшает его выведение из плазмы, поэтому он имеет относительно высокое время полувыведения — от 60 до 90 минут. Связанные с белками плазмы крови кортикостероиды играют роль резерва, препятствующего быстрым колебаниям концентрации свободных гормонов в крови, что происходит, например, с кортизолом в связи с кратковременным стрессом и эпизодической продукцией АКТГ. Этот функциональный резерв может также обеспечивать равномерное распределение гормонов по тканям.
Некоторые глюкокортикоиды обладают как глюкокортикоидной, так и минералокортикоидной активностью. Кортизол, обладающий слабой минералокортикоидной активностью (1/30 активности альдостерона), ввиду громадной его продукции при некоторых синдромах может вызывать минералокортикоидные ответы наряду с более выраженными глюкокортикоидными влияниями. Синтетические глюкокортикоиды различаются по своей активности относительно кортизола: преднизон в 4 раза активнее кортизола, метилпреднизон в 5 раз активнее преднизона, а дексаметазон в 30 раз активнее кортизона. Дексаметазон является синтетическим препаратом с нулевой минералокортикоидной и чрезвычайно высокой глюкокортикоидной активностью, что имеет важное значение для получения чистых глюкокортикоидных ответов без побочных эффектов задержки натрия и воды.
Альдостерон — ключевой регулятор натриевого и калиевого обмена
Альдостерон является ключевым гормоном, регулирующим натриевый и калиевый обмен в эпителиальных тканях организма. В почечных канальцах гормон усиливает реабсорбцию натрия и одновременно повышает секрецию калия, активируя натрий-калиевую АТФ-азу на базолатеральной мембране эпителиоцитов. При этом альдостерон также увеличивает проницаемость апикальной мембраны для натрия, что облегчает его поступление из просвета канальца в клетку. Координация этих процессов обеспечивает эффективный транспорт ионов по обе стороны эпителиальной клетки: натрий проходит из канальца в кровь, а калий выводится в просвет, где далее удаляется с мочой.
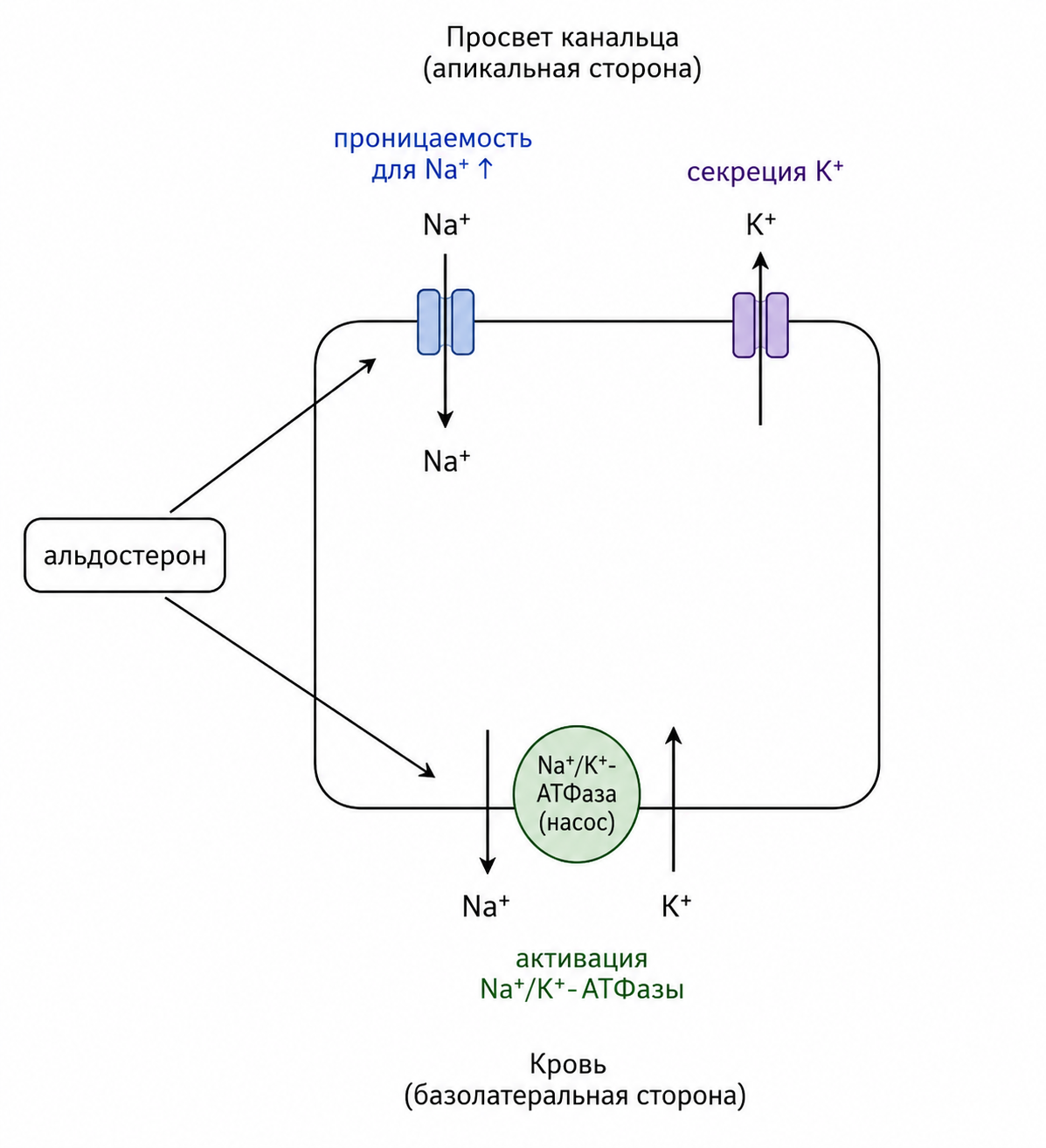 Рис. 2. Схема действия альдостерона на эпителиоцит почечного канальца.
Рис. 2. Схема действия альдостерона на эпителиоцит почечного канальца.
На клеточном уровне альдостерон действует преимущественно геномным механизмом. После связывания с внутриклеточным рецептором гормон инициирует серию транскрипционных событий, приводящих к синтезу специфических белков-переносчиков натрия. Появление новой мРНК занимает около тридцати минут, увеличение транспортной активности достигает своего пика лишь примерно через сорок пять минут, а максимальный эффект наблюдается только через несколько часов. Такой замедленный характер ответа указывает на то, что основное действие альдостерона реализуется через изменение генетической программы клетки.
Тем не менее, помимо медленного геномного пути, альдостерон способен вызывать быстрые негеномные реакции, проявляющиеся в течение секунд-минут. Эти эффекты, вероятно, опосредованы взаимодействием гормона с рецепторами, принадлежащими клеточным мембранам и связанными с системами вторичных посредников. В течение менее чем двух минут альдостерон усиливает синтез цАМФ в эпителиоцитах почечных канальцев и в гладкомышечных клетках сосудов, а в некоторых типах клеток быстро активирует фосфатидилинозитольные сигнальные пути. Структуры рецепторов, отвечающих за такие быстрые ответы, пока не определены, и их физиологическое значение остаётся предметом исследований.
Отсутствие альдостерона, как при болезни Аддисона, приводит к потере натрия, удержанию калия и усиленному выводу воды, что уменьшает объём внеклеточной жидкости и понижает артериальное давление. При полном дефиците гормона снижение объёма жидкости может стать выраженным, особенно при ограничении потребления соли и воды с пищей. Противоположная ситуация наблюдается при избыточной секреции альдостерона, например при синдроме Конна: задерживается натрий, а запасы калия истощаются. Несмотря на то, что небольшие количества альдостерона достаточны для регулирования натрия, его роль в контроле калиевого баланса гораздо значительнее, так как именно он определяет степень калиевой экскреции с мочой.
Регуляция продукции альдостерона тесно связана с изменениями концентрации электролитов во внеклеточной жидкости, объёма крови, артериального давления и функций почек, а также с действием ангиотензина II — самого мощного натрийудерживающего гормона, уровень которого растёт при гипотонии и гиповолемии. Помимо почек, альдостерон воздействует на эпителиальные ткани потовых и слюнных желёз, а также на кишечный эпителий, где аналогично регулирует натриевую реабсорбцию и калиевую секрецию. Натрий, попадая через апикальную мембрану в клетку, затем перекачивается Na⁺/K⁺-насосом, расположенным в базолатеральной мембране; альдостерон усиливает оба этих этапа, обеспечивая эффективный перенос натрия из просвета канальца в межклеточное пространство и далее в кровоток.
Мозговое вещество: катехоламины и реакция «борьбы-бегства»
В мозговом веществе надпочечников тирозин проходит через последовательные стадии гидроксилирования и декарбоксилирования, сначала образуя дофамин, затем норадреналин; окончательная метиляция норадреналина ферментом фенилэтиламин-N-метилтрансферазой превращает около 80 % этого катехоламина в адреналин. Этим объясняется высокий процент адреналина в крови по сравнению с норадреналином и отличие надпочечникового мозгового вещества от симпатических нервных окончаний, где метилирование происходит в значительно меньших количествах.
При стрессовых или тревожных стимулах гипоталамус активирует ретикулярную формацию ствола мозга, откуда импульсы спускаются по симпатическому тракту к надпочечникам; ацетилхолин, высвобождённый из симпатических предшественников, стимулирует хромаффинные клетки мозгового вещества, ускоряя синтез и секрецию адреналина и норадреналина. Это мгновенно усиливает гормональную поддержку реакции «борьбы-или-бегства», позволяя симпатической нервной системе одновременно повышать кровообращение в скелетных мышцах, артериальное давление и концентрацию глюкозы в плазме, ускоряя гликолиз в печени и мышцах. Адреналин и норадреналин, попадая в кровь, связываются с α- и β-адренорецепторами на клетках сердца, сосудов, печени и скелетных мышц, вызывая усиление силы мышечных сокращений и повышение умственной активности в условиях острого стресса.
После высвобождения норадреналин из нервных окончаний удаляется из синаптической щели тремя путями: активным обратным захватом адренергическими нейронами (удаляется 50–80 % медиатора), диффузией в окружающие ткани с последующим попаданием в кровь, что выводит большую часть оставшегося норадреналина, и ферментативным разрушением небольшого количества медиатора тканевыми ферментами, включая моноаминоксидазу в нервных окончаниях и катехол-О-метилтрансферазу, распределённую во всех тканях. Норадреналин, не попадающий непосредственно в ткань, сохраняет биологическую активность лишь несколько секунд, поскольку быстро захватывается обратно или диффундирует; в отличие от него, адреналин и норадреналин, секретируемые мозговым веществом в кровь, остаются активными 10–30 секунд, после чего их действие постепенно угасает в течение от одной до нескольких минут, пока они не диффундируют в ткани, где их разрушает катехол-О-метилтрансфераза, преимущественно в печени.
Для стимуляции эффекторных органов катехоламины обязаны связываться со специфическими рецепторами, расположенными в наружной части клеточной мембраны. Эти рецепторы состоят из внешней части, взаимодействующей с медиатором, и внутренней трансмембранной домены, которая при конформационном изменении активирует или инактивирует ионные каналы либо ферменты, вызывая возбуждение или торможение клетки-эффектора.
В печени катехол-О-метилтрансфераза метилирует адреналин и норадреналин, превращая их в менее активные метаболиты; благодаря этому их физиологический эффект полностью исчезает в течение нескольких минут после стимуляции, обеспечивая быстрый и контролируемый отклик симпатической стрессовой реакции. Такая скоростная инактивация критически важна: без неё однократный эпизод стресса мог бы перерасти в длительную катехоламиновую интоксикацию.
Адреналовые андрогены и их периферическая конверсия
Адреналовые андрогены представляют собой умеренно активные мужские половые гормоны, секретируемые корой надпочечников на протяжении всей жизни, включая внутриутробный период развития. Наиболее значимым из них является дегидроэпиандростерон (ДГЭА); кроме того, в коре надпочечников в минимальных количествах продуцируются прогестерон и эстрогены, что отражает гормональную универсальность этого органа. В норме эти гормоны оказывают лишь слабое влияние на организм, однако их постоянное присутствие в крови обеспечивает фоновое андрогенное воздействие на ткани-мишени, что становится клинически значимым при патологических состояниях.
Уже с предпубертатного периода адреналовые андрогены способны влиять на женский организм, проявляясь, в частности, в росте волос подмышечной области и на лобке. Это демонстрирует их роль в формировании вторичных половых признаков и подчёркивает многогранность физиологического действия этих гормонов у женщин на протяжении всей жизни.
Некоторые адреналовые андрогены подвергаются периферической конверсии во внеадреналовых тканях, где превращаются в тестостерон — основной мужской половой гормон с наибольшей биологической активностью. Таким образом, они служат предшественниками для синтеза более активных андрогенов, что существенно усиливает их физиологическое и клиническое значение. При патологии коры надпочечников, когда синтезируются чрезмерно большие количества андрогенов, их избыток приводит к маскулинизации организма, указывая на то, что даже умеренно активные гормоны при перепроизводстве способны вызывать выраженные клинические эффекты гиперандрогении.
Пучковая зона коры надпочечников, составляющая около 75 % её объёма и являющаяся наиболее широкой, наряду с синтезом глюкокортикоидов (кортизол и кортикостерон) продуцирует также небольшое количество андрогенов и эстрогенов. Это означает, что андрогены, как и глюкокортикоиды, являются продуктами одной и той же зоны коры, их секреция регулируется АКТГ гипофиза, а нарушение функции пучковой зоны может одновременно затрагивать несколько гормональных осей.
Тестостерон, образованный из адренальных андрогенов в периферических тканях, способен стимулировать рост волос на различных участках тела, усиливая этот эффект на кожных областях, чувствительных к андрогенному действию. Этот механизм иллюстрирует, как даже умеренно активные андрогены надпочечников, превращаясь в тестостерон, могут влиять на пилозный покров и обеспечивать физиологические эффекты, характерные для мужских половых гормонов.
Физиологические эффекты андрогенов традиционно обсуждаются в контексте мужских половых функций, однако адреналовые андрогены обладают собственным спектром действия, проявляющимся как в мужском, так и в женском организме. Их влияние на оволосение, гормональный баланс и потенциальные эффекты маскулинизации подчёркивает значимость надпочечниковых андрогенов как самостоятельного фактора эндокринной регуляции, а не только как предшественников тестостерона.
Регуляция оси гипоталамус–гипофиз–надпочечники
Кортикотропин-рилизинг-гормон (КРГ) представляет собой пептид из 41 аминокислоты, синтезируемый и накапливающийся в специализированных нейронах парвентрикулярного ядра гипоталамуса; после высвобождения он поступает в гипофизарный портал, где стимулирует продукцию адренокортикотропного гормона (АКТГ) в передней доле гипофиза. Отсутствие КРГ приводит к резкому снижению АКТГ, подтверждая его ключевую роль в инициации гормональной цепи.
АКТГ достигает адренокортикальных клеток надпочечников, связываясь с рецептором, сопряжённым с аденилатциклазой, что вызывает быстрый рост внутриклеточного уровня циклического аденозинмонофосфата (цАМФ); пиковое увеличение цАМФ наблюдается примерно через три минуты после стимула и запускает ферментативные реакции, необходимые для синтеза кортизола. Стимуляция АКТГ активирует протеинкиназу А, которая катализирует первую, решающую стадию превращения холестерола в прегненолон; этот шаг определяет лимитирующий поток в стероидогенезе, позволяя последовательно образовывать кортизол и другие надпочечные стероиды.
Концентрация кортизола в плазме подчиняется чётко выраженному суточному ритму: после пробуждения наблюдается утренний всплеск в течение первого часа, затем уровень постепенно снижается до минимума к полудню и достигает наименьшего значения в ночное время. Этот циркадный паттерн обусловлен ритмичной секрецией КРГ и, следовательно, АКТГ, что превращает надпочечники в часы, синхронизированные с циклом «сон–бодрствование».
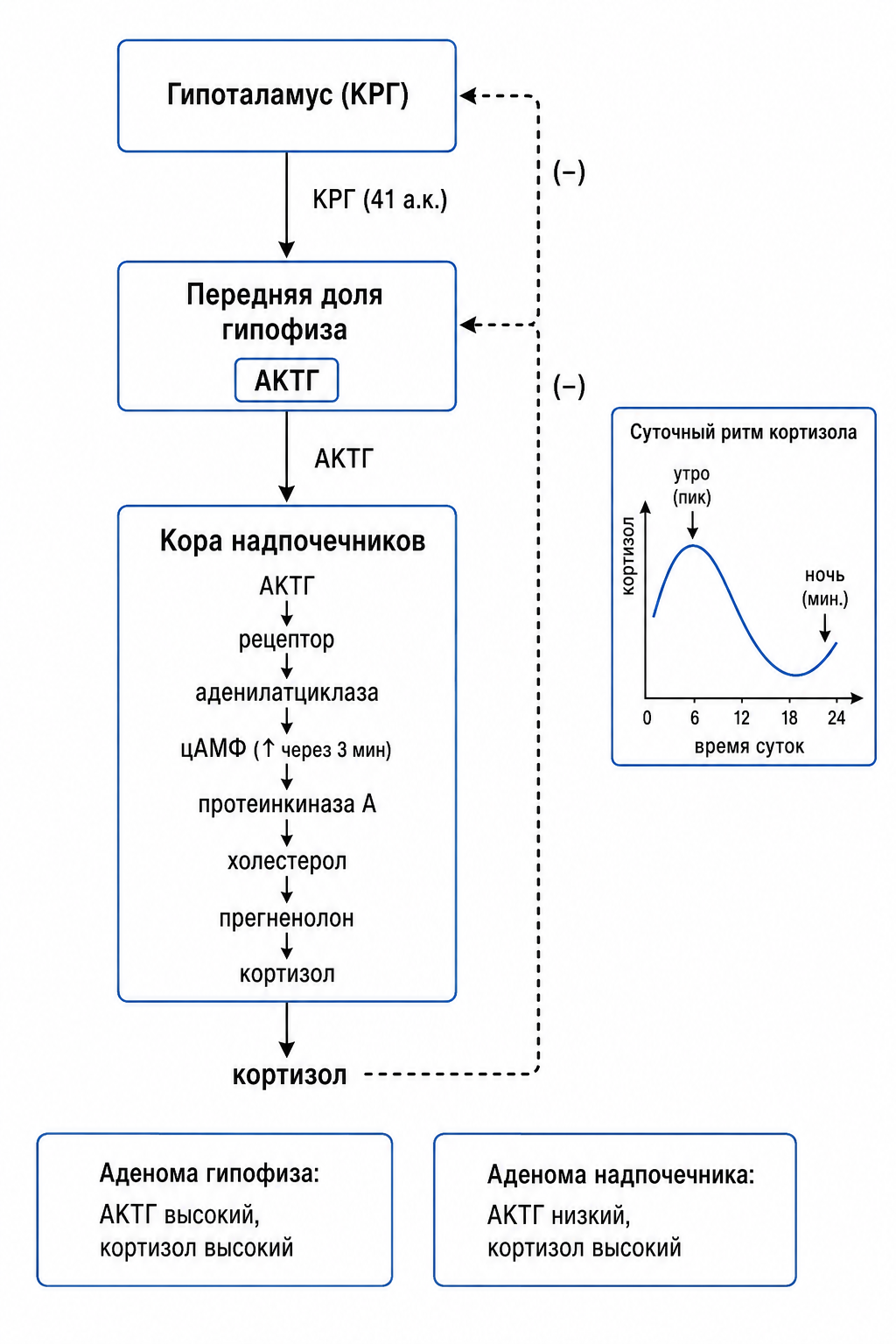 Рис. 3. Схема оси гипоталамус-гипофиз-надпочечники.
Рис. 3. Схема оси гипоталамус-гипофиз-надпочечники.
Отрицательная обратная связь регулирует ось гипоталамус–гипофиз–надпочечники: повышенные плазменные концентрации кортизола подавляют синтез и высвобождение КРГ из гипоталамуса и АКТГ из гипофиза, тем самым ограничивая дальнейшую продукцию кортизола. При хронически повышенных уровнях кортизола, например при аденоме гипофиза, уровень АКТГ остаётся высоким, тогда как при первичной гиперсекреции кортизола (аденоме коры надпочечников) уровень АКТГ снижается в результате подавления обратной связи. Таким образом, измерение АКТГ позволяет клинически различить центральный и периферический источник гиперкортицизма.
Патологии надпочечников
При болезни Аддисона, характеризующейся разрушением коры надпочечников, отсутствует ответ на стресс-индуцированную секрецию кортикостероидов; любой физический или психический стресс приводит к аддисоническому кризу, проявляющемуся острой недостаточностью глюкокортикоидов и требующим немедленной заместительной терапии. Без введения кортизола или аналогов такое состояние может закончиться летально в течение часов.
Синдром Кушинга возникает при гиперсекреции кортизола, часто следствием избыточного выхода АКТГ; наиболее частая этиология — аденома передней доли гипофиза, продуцирующая избыточный АКТГ, что приводит к гипертрофии надпочечников и повышенному выведению кортизола. В 20–25 % клинических случаев избыток кортизола обусловлен первичной гиперпродукцией коры надпочечников, при этом уровень АКТГ снижается из-за подавления отрицательной обратной связи. Клиническая картина включает характерное перераспределение жира, артериальную гипертензию, мышечную слабость и нарушения углеводного обмена.
Заключение
Надпочечники представляют собой уникальный эндокринный орган, в котором тесно переплетены кора, синтезирующая стероидные гормоны, и мозговое вещество, продуцирующее катехоламины. Каждая из трёх зон коры — клубочковая, пучковая и сетчатая — выполняет строго определённую функцию, обеспечивая организм минералокортикоидами, глюкокортикоидами и андрогенами соответственно. Лимитирующая стадия стероидогенеза, превращение холестерола в прегненолон под действием цитохрома P450scc, становится точкой интеграции сигналов АКТГ и ангиотензина II, что позволяет одной и той же биохимической реакции порождать совершенно разные гормональные ответы.
Кортизол, обеспечивающий около 95 % глюкокортикоидной активности, управляет глюконеогенезом, мобилизацией жирных кислот и противовоспалительной защитой, в то время как альдостерон, обладая в тысячи раз более низкой плазменной концентрацией, благодаря 3000-кратному аффинитету к рецепторам обеспечивает почти 90 % минералокортикоидной активности. Мозговое вещество дополняет эту систему мгновенной катехоламиновой реакцией «борьбы-бегства», а адреналовые андрогены формируют фоновый половой фон, становящийся клинически значимым при патологии. Регуляция всей этой архитектуры осуществляется осью гипоталамус–гипофиз–надпочечники с циркадным ритмом и отрицательной обратной связью, а её нарушения — при болезни Аддисона и синдроме Кушинга — наглядно демонстрируют, насколько критически важен этот небольшой орган для поддержания гомеостаза, адаптации к стрессу и самой возможности жизни.
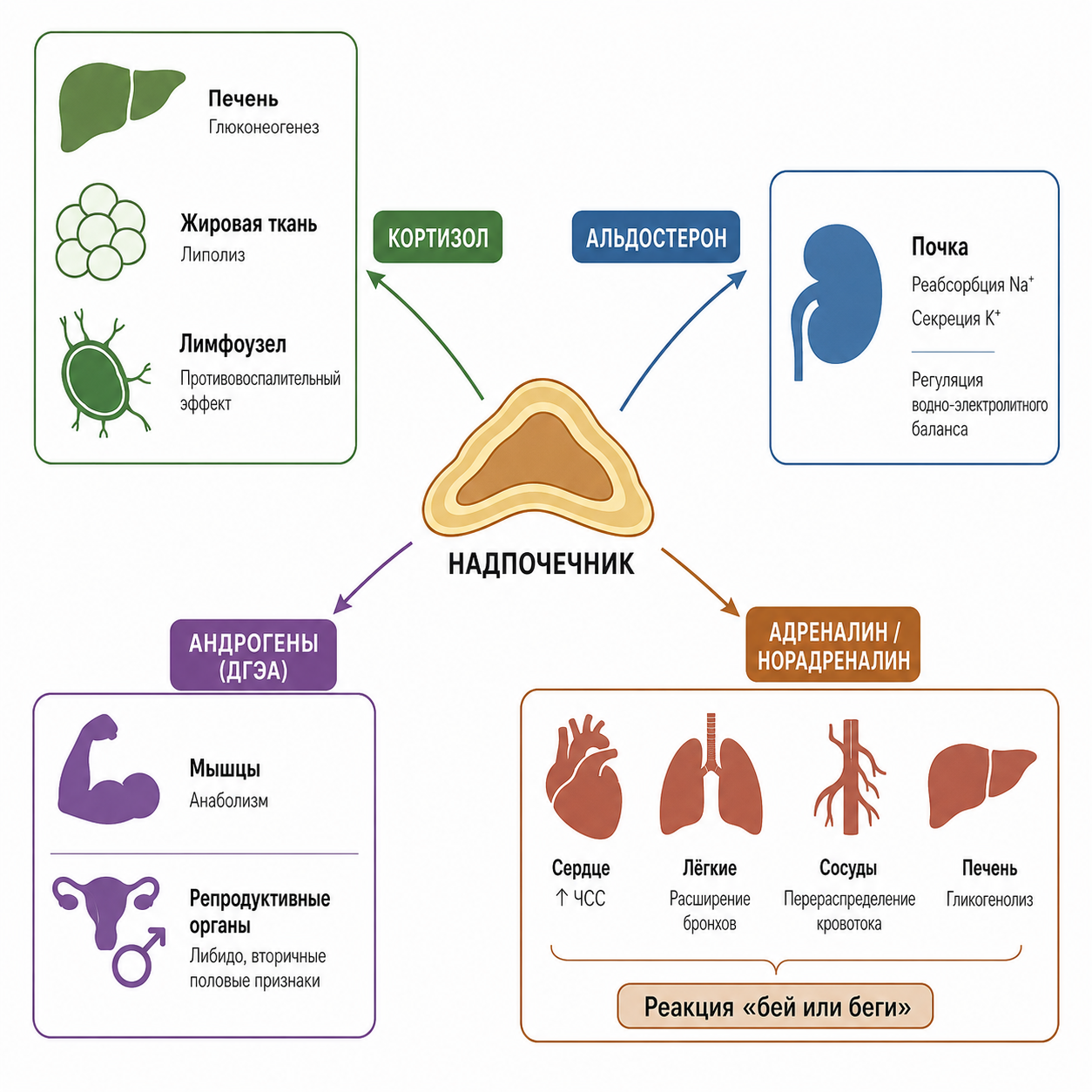 Рис. 4. Суммирующая схема о роли надпочечников.
Рис. 4. Суммирующая схема о роли надпочечников.