Строение растительной клетки
Общая характеристика и отличительные особенности
Растительная клетка является эукариотической: она обладает морфологически оформленным ядром и сложной системой внутренних мембран. От клеток животных её отличают три обязательных признака — жёсткая целлюлозная клеточная стенка, специфические органеллы (пластиды) и крупная центральная вакуоль, заполненная клеточным соком.
Живое содержимое клетки называют протопластом; от внешней среды или клеточной стенки его отделяет плазматическая мембрана. Протопласт состоит из ядра и цитоплазмы, в которой распределены органеллы и системы мембран.
 Рис. 1. Особенности растительной клетки
Рис. 1. Особенности растительной клетки
Плазматическая мембрана (плазмалемма)
Плазмалемма представляет собой одиночную мембрану, ограничивающую цитоплазму. На электронных микрофотографиях она выглядит как трёхслойная структура («элементарная мембрана»). Мембрана регулирует транспорт веществ в протопласт и из него, координирует синтез целлюлозных микрофибрилл клеточной стенки, а также передаёт сигналы от гормонов и окружающей среды, контролируя рост и дифференцировку. В её состав входят белки-каналы аквапорины, облегчающие и ускоряющие ток воды, что критически важно для поддержания осмотического равновесия между вакуолью и цитозолем.
Ядро
Ядро ограничено ядерной оболочкой, состоящей из двух липопротеидных мембран, пронизанных порами для связи с цитоплазмой. Внутри находится нуклеоплазма (основное вещество), в которой распределены хроматин (комплекс ДНК и белков-гистонов) и одно или несколько ядрышек.
Функции ядра — сохранение и передача генетической информации, контроль метаболизма и упорядоченное развитие клетки и её потомков. Здесь происходят процессы репликации и транскрипции. Во время деления клетки (митоза) ядро претерпевает значительные изменения, а при дифференцировке некоторых специализированных клеток, например члеников ситовидных трубок, оно может разрушаться (пикнотическая дегенерация).
Цитоплазма и цитозоль
Цитоплазма включает все компоненты клетки, кроме ядра. Её матрикс называют цитозолем (или гиалоплазмой). Это сложный водный раствор, в котором находятся все органеллы и протекают многие биохимические реакции, в том числе гликолиз. Цитозоль пронизан сетью цитоскелета (микротрубочки и актиновые филаменты), который обеспечивает перемещение органелл и движение самой цитоплазмы — циклоз.
Клеточная стенка и плазмодесмы
Общая характеристика и функции
Клеточная стенка (оболочка) — жёсткий внеклеточный каркас, ограничивающий размер протопласта, определяющий форму клетки и защищающий её от механических повреждений. Стенка метаболически активна и участвует в поглощении, транспорте и секреции веществ. Её наличие является главным отличием растений от животных.
Химический состав
Основной компонент — целлюлоза, линейные цепи полисахарида глюкана. Молекулы целлюлозы объединяются в микрофибриллы, прочность на разрыв которых близка к стали. Микрофибриллы погружены в матрикс, состоящий из гемицеллюлоз (например, ксилоглюканов, связывающих микрофибриллы и ограничивающих растяжимость стенки), пектиновых веществ (гидрофильных полисахаридов, образующих гель и определяющих пористость и пластические свойства) и белков-гликопротеинов, таких как экстенсины, придающие стенке прочность.
В стенку могут включаться инкрустирующие вещества: лигнин вызывает одревеснение, придавая жёсткость и водонепроницаемость (характерно для древесины), а суберин и кутин — жироподобные вещества, обеспечивающие защиту от потери воды (пробка, кутикула).
 Рис. 2. Состав и слои клеточной стенки. Middle Lamella - срединная пластинка; Primary Cell Wall - первичная клеточная стенка; Plasma Membrane - плазматическая мембрана; Pectin - пектин; Cellulose Microfibril - целлюлозные микрофибриллы; Hemicellulose - гемицеллюлоза
Рис. 2. Состав и слои клеточной стенки. Middle Lamella - срединная пластинка; Primary Cell Wall - первичная клеточная стенка; Plasma Membrane - плазматическая мембрана; Pectin - пектин; Cellulose Microfibril - целлюлозные микрофибриллы; Hemicellulose - гемицеллюлоза
Слои клеточной стенки
Стенка формируется протопластом центробежно (от внешней стороны внутрь), поэтому самый молодой слой всегда находится ближе всего к мембране.
Срединная пластинка — самый наружный слой, общий для двух соседних клеток; состоит преимущественно из пектина и склеивает клетки между собой.
Первичная клеточная стенка формируется, пока клетка растёт. Микрофибриллы целлюлозы в ней расположены рыхло и беспорядочно, что позволяет стенке растягиваться.
Вторичная клеточная стенка откладывается изнутри первичной после прекращения роста. Она богаче целлюлозой и часто состоит из трёх слоёв (S1, S2, S3), различающихся ориентацией микрофибрилл.
Поры и поровые поля
Первичные поровые поля — тонкие участки первичной стенки, через которые проходят группы плазмодесм. Поры — углубления (разрывы) во вторичной стенке, возникающие над первичными поровыми полями. Поры соседних клеток обычно располагаются друг против друга, образуя пару пор, разделённую поровой мембраной.
Плазмодесмы, симпласт и апопласт
Плазмодесмы — тонкие тяжи цитоплазмы, пронизывающие стенки и соединяющие соседние протопласты. Канал плазмодесмы выстлан мембраной, а внутри него проходит десмотрубочка — видоизменённая структура эндоплазматического ретикулума. Плазмодесмы обеспечивают транспорт воды, ионов, сахаров и даже макромолекул (белков, вирусов) между клетками. Благодаря им все живые протопласты растения объединяются в единую систему — симпласт. Систему клеточных стенок и межклетников называют апопластом.
Формирование стенки (цитокинез)
Новая стенка закладывается в конце деления ядра. Образуется фрагмопласт (система микротрубочек и актиновых филаментов), который направляет везикулы аппарата Гольджи к плоскости деления. Слияние везикул формирует клеточную пластинку, растущую от центра к периферии, пока она не соединится со стенками материнской клетки.
Пластиды
Общая характеристика и происхождение
Пластиды, наряду с клеточной стенкой и вакуолями, являются обязательными компонентами растительных клеток. Это полуавтономные органеллы; согласно теории симбиогенеза, они произошли от свободноживущих цианобактерий путём эндосимбиоза. В пользу этого свидетельствуют структурные особенности пластид, сходные с бактериальными: наличие собственного генома в виде кольцевой ДНК, не связанной с гистонами; собственные рибосомы 70S-типа (в цитоплазме эукариот — 80S); способность к размножению бинарным делением независимо от деления ядра; оболочка из двух мембран, где внутренняя мембрана по составу схожа с бактериальной.
Внутреннее строение
Внутренняя среда пластиды называется стромой (полужидкий матрикс). В ней расположена система внутренних мембран — тилакоидов. Тилакоиды могут быть собраны в стопки — граны, которые соединены между собой одиночными стромальными тилакоидами.
Классификация по типу пигментов
Хлоропласты — зелёные пластиды, содержащие хлорофиллы и каротиноиды. В них происходит фотосинтез, то есть преобразование световой энергии в химическую энергию углеводов. Хлоропласты способны перемещаться в клетке, оптимизируя поглощение света: при слабом свете они выстраиваются параллельно поверхности листа, при слишком ярком — перпендикулярно, чтобы избежать повреждений.
 Рис. 3. Строение хлоропласта. 1 - ламелла; 2 - межмембранное пространство; 3 - наружняя мембрана; 4 - внутренняя мембрана; 5 - строма; 6 - тилакоид; 7 - грана; 8 - липидные капли; 9 - ДНК; 10 - люмен; 12 - крахмальное зерно
Рис. 3. Строение хлоропласта. 1 - ламелла; 2 - межмембранное пространство; 3 - наружняя мембрана; 4 - внутренняя мембрана; 5 - строма; 6 - тилакоид; 7 - грана; 8 - липидные капли; 9 - ДНК; 10 - люмен; 12 - крахмальное зерно
Kārlis Kalviškis, CC BY-SA 4.0 https://creativecommons.org/licenses/by-sa/4.0, via Wikimedia Commons
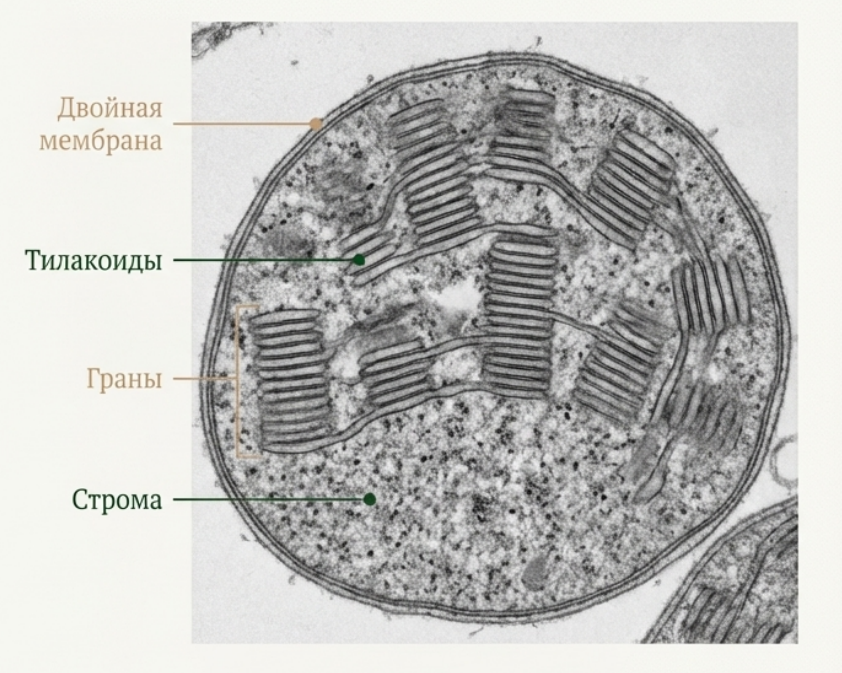 Рис. 4. Фото хлоропласта под электронным микроскопом.
Рис. 4. Фото хлоропласта под электронным микроскопом.
Хромопласты — пигментированные пластиды жёлтого, оранжевого или красного цвета, содержащие только каротиноиды. Они определяют яркую окраску лепестков, плодов и осенних листьев, привлекая опылителей и распространителей семян. Хромопласты могут развиваться как из пропластид, так и из зрелых хлоропластов (например, при созревании томатов).
Лейкопласты — бесцветные пластиды, специализирующиеся на синтезе и хранении веществ. К ним относятся:
амилопласты — накапливают запасной крахмал в виде зёрен; крахмальные зёрна имеют слоистую структуру и в поляризованном свете дают характерный эффект «мальтийского креста»;
элайопласты (олеопласты) — синтезируют и запасают масла;
протеинопласты — накапливают белки.
Развитие и взаимопревращения
Все пластиды происходят от пропластид — мелких бесцветных органелл, находящихся в меристемах (делящихся тканях). Пластиды обладают высокой пластичностью: в темноте пропластиды превращаются в этиопласты, содержащие проламеллярные тела (кристаллические решётки из мембран); на свету эти структуры быстро преобразуются в тилакоиды хлоропластов. Амилопласты в клубнях картофеля на свету могут зеленеть, превращаясь в хлоропласты.
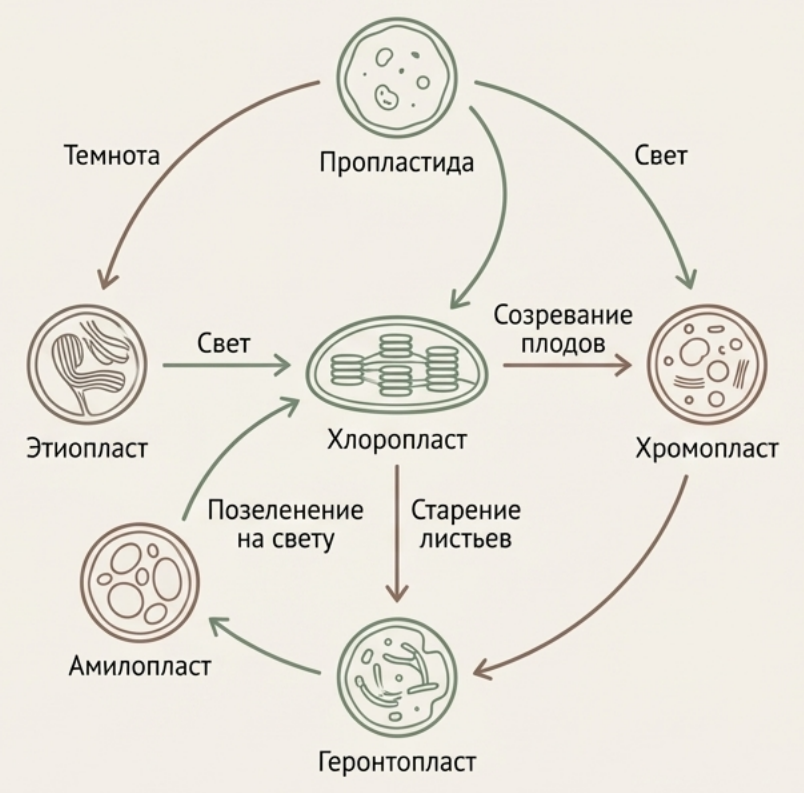 Рис. 5. Взаимопревращения пластид
Рис. 5. Взаимопревращения пластид
Вакуоли
Вакуоль — один из трёх главных отличительных признаков растительной клетки. Она ограничена одной мембраной — тонопластом (вакуолярной мембраной). Внутри находится клеточный сок — водный раствор, состав которого варьируется в зависимости от типа клетки и стадии её развития.
В молодых меристематических клетках вакуолей много и они мелкие. По мере роста клетки они сливаются в одну крупную центральную вакуоль, способную занимать до 90% объёма зрелой клетки, оттесняя цитоплазму к клеточной стенке. Функции вакуоли: поддержание тургора — накопление осмотически активных веществ (сахаров, солей) заставляет воду поступать в вакуоль, что создаёт давление на клеточную стенку и придаёт тканям упругость; запасание первичных метаболитов (сахаров, органических кислот) и белков (особенно в семенах); защита и детоксикация — вакуоль изолирует токсичные вторичные метаболиты (алкалоиды, таннины), защищающие растение от поедания; окраска — водорастворимые пигменты антоцианы в вакуолях придают красную, синюю и фиолетовую окраску цветам и плодам; литическая функция — некоторые вакуоли содержат гидролитические ферменты для расщепления макромолекул и переработки старых органелл, выполняя роль лизосом.
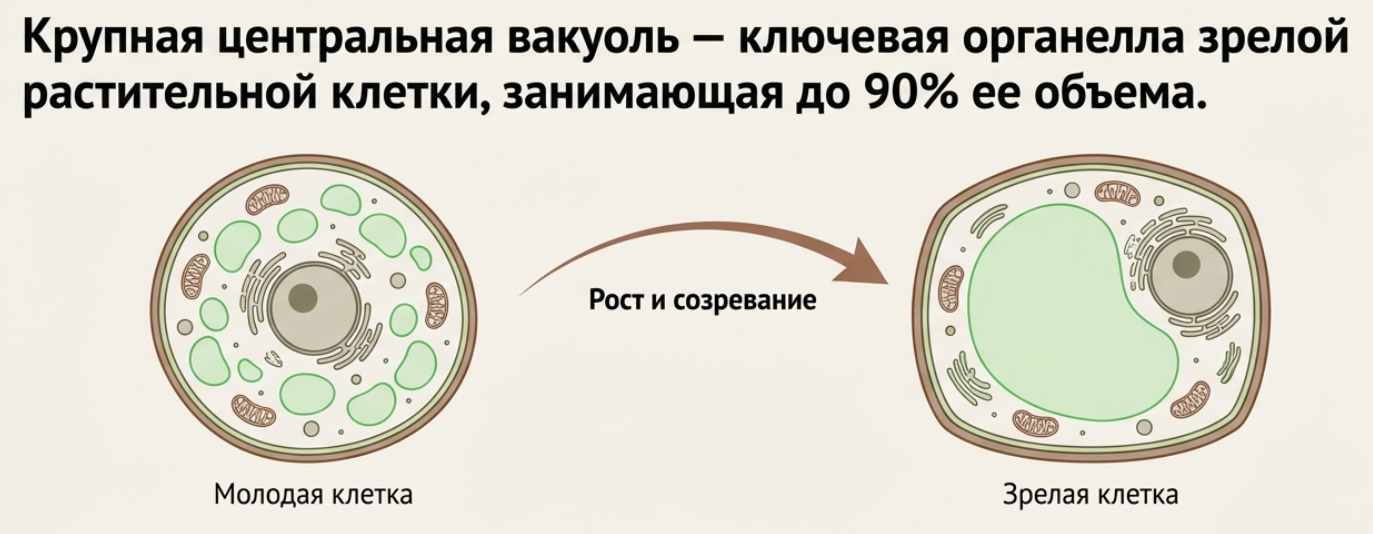 Рис. 6. Рост вакуоли при созревании растительной клетки
Рис. 6. Рост вакуоли при созревании растительной клетки
Митохондрии
Митохондрии обеспечивают клетку энергией в форме АТФ путём дыхания. Они окружены оболочкой из двух мембран. Внутренняя мембрана образует многочисленные складки — кристы, увеличивающие рабочую площадь для ферментов. Внутреннее пространство заполнено матриксом, содержащим ферменты цикла лимонной кислоты, воду и фосфаты.
Как и пластиды, митохондрии полуавтономны: они содержат собственный геном (кольцевую ДНК, не связанную с гистонами) и собственные рибосомы 70S-типа, способны к самостоятельному делению и слиянию. Согласно теории симбиогенеза, митохондрии произошли от свободноживущих α-протеобактерий, вступивших в симбиоз с древней клеткой-хозяином. Митохондрии постоянно перемещаются в цитоплазме под действием актин-миозинового комплекса. В фотосинтезирующих клетках они часто располагаются в тесном контакте с хлоропластами и пероксисомами, что необходимо для координации процессов фотодыхания и эффективного обмена энергией.
Система внутренних мембран
Эндоплазматический ретикулум (ЭР) ЭР представляет собой непрерывную трёхмерную систему мембран, пронизывающую весь цитозоль. В разрезе он выглядит как две параллельные мембраны с узким пространством между ними — люменом. Внешняя мембрана ядерной оболочки напрямую соединяется с ЭР, делая его продолжением ядерного компартмента.
Различают два типа ЭР. Шероховатый (гранулярный) состоит из плоских мешочков (цистерн), усыпанных рибосомами; здесь происходит синтез белков, предназначенных для секреции или встраивания в мембраны. Гладкий (агранулярный) имеет преимущественно трубчатую форму и лишён рибосом; он специализируется на синтезе липидов (например, мембранных масел и восков). Мембраны ЭР находятся в постоянном движении, меняя форму и расположение под действием цитоскелета.
Аппарат (комплекс) Гольджи
Это сильно поляризованная мембранная система, состоящая из отдельных стопок дисковидных цистерн — диктиосом. Каждая стопка имеет цис-сторону (принимающую) и транс-сторону (отправляющую). Везикулы отпочковываются от ЭР и сливаются с аппаратом Гольджи, где содержимое модифицируется. Главная роль аппарата Гольджи в растительной клетке — синтез нецеллюлозных полисахаридов (пектинов и гемицеллюлоз) для клеточной стенки. Эти вещества упаковываются в везикулы и выводятся наружу путём экзоцитоза. Здесь же происходит гликозилирование — присоединение к белкам углеводных цепочек с образованием гликопротеинов.
Цитоскелет
Цитоскелет — динамичная сеть белковых нитей, пронизывающая цитозоль. В его составе выделяют два основных типа структур.
Микротрубочки — полые цилиндры из белка тубулина. Для них характерна «динамическая неустойчивость»: они постоянно собираются и разбираются. Микротрубочки управляют ориентацией целлюлозных волокон при строительстве клеточной стенки, формируют веретено деления и фрагмопласт при цитокинезе.
Актиновые филаменты (микрофиламенты) — тонкие спиральные нити из белка актина. Они обеспечивают движение самой цитоплазмы (циклоз), перемещение органелл и везикул, а также определяют полярность клетки при росте (например, у корневых волосков).
Рибосомы
Рибосомы — мелкие частицы, состоящие из РНК и белка, на которых происходит сборка аминокислот в белковые цепи. В растительной клетке они могут быть свободными в цитозоле, прикреплёнными к ЭР и ядерной оболочке, а также находиться внутри полуавтономных органелл (пластид и митохондрий).
Эргастические вещества и клеточные включения
Эргастические вещества — пассивные продукты жизнедеятельности протопласта; в отличие от органелл, они не являются живыми компонентами клетки. Эти вещества могут появляться и исчезать на разных стадиях жизни клетки и локализоваться в цитозоле, вакуолях, пластидах или встраиваться в клеточную стенку.
Запасные углеводы (крахмал)
Крахмал — самый распространённый запасной полисахарид растений. Ассимиляционный крахмал образуется непосредственно в хлоропластах в процессе фотосинтеза в виде мелких временных зёрен. Запасной крахмал синтезируется в амилопластах из сахаров, притекающих из листьев. Крахмальные зёрна имеют слоистую структуру, отражающую суточные ритмы синтеза; слои располагаются вокруг образовательного центра — хилума. Амилопласты могут быть простыми (одно зерно, как у картофеля) или сложными (множество мелких зёрнышек в одной оболочке, как у риса или овса). В поляризованном свете крахмальные зёрна дают характерный эффект «мальтийского креста».
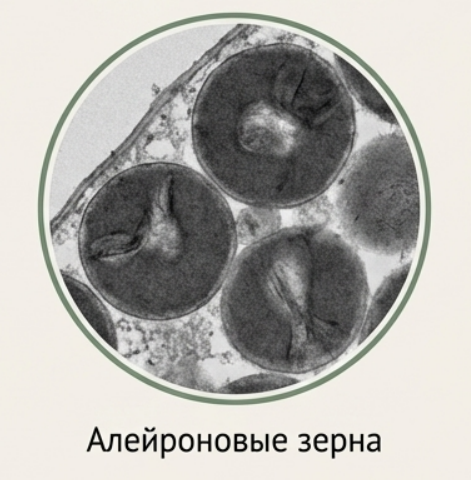
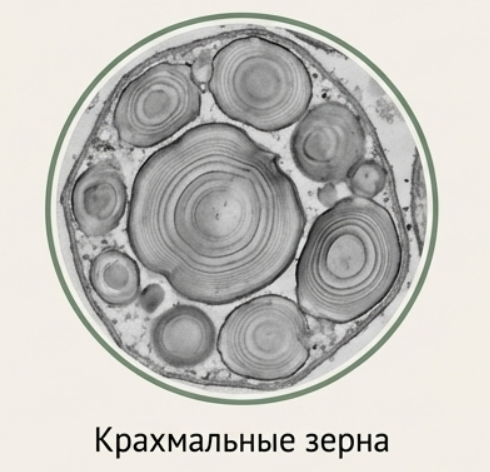 ## Запасные белки (алейроновые зёрна)
## Запасные белки (алейроновые зёрна)
Белки запасаются преимущественно в семенах в виде белковых телец, или алейроновых зёрен. При созревании семян вакуоли теряют воду, и белки выпадают в осадок, превращаясь в твёрдые зёрна. Сложные алейроновые зёрна (например, у клещевины) могут содержать включения: кристаллоид (белковый кристалл) и глобоид (скопление солей фитиновой кислоты, богатых фосфором).
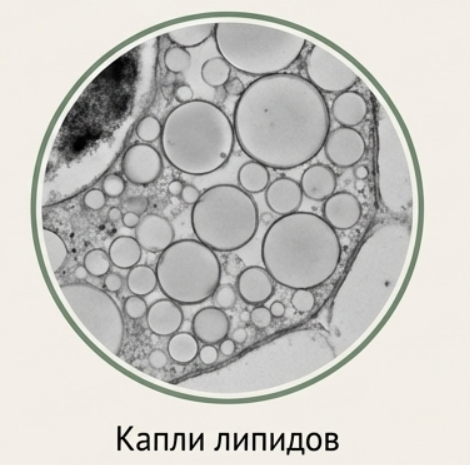
 ## Запасные жиры (масляные тельца)
## Запасные жиры (масляные тельца)
Жиры — наиболее калорийное запасное вещество, встречающееся почти у 90% видов растений. Масляные тельца (олеосомы) — сферические структуры в цитоплазме, выглядящие на электронных микрофотографиях аморфными. Они образуются при участии эндоплазматического ретикулума. Особенно богаты жирами семена подсолнечника, рапса и плоды масличных культур.
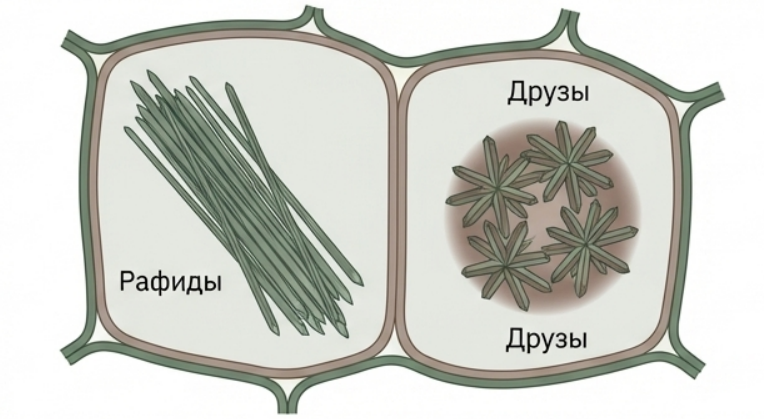
 ## Минеральные включения (кристаллы)
## Минеральные включения (кристаллы)
Кристаллы являются конечными продуктами обмена веществ и чаще всего образуются в вакуолях. Наиболее частый компонент — оксалат кальция, имеющий несколько форм: рафиды (игловидные кристаллы, собранные в плотные пучки — характерны для винограда, бальзамина), друзы (сферические звёздчатые сростки), стилоиды (одиночные крупные удлинённые кристаллы), кристаллический песок (масса очень мелких кристаллов). Карбонат кальция образует цистолиты — крупные гроздевидные образования, возникающие в специализированных клетках-литоцистах. Цистолит обычно прикреплён к клеточной стенке на специальном стебельке.
 ## Таннины (дубильные вещества)
## Таннины (дубильные вещества)
Таннины — группа фенольных соединений, накапливающихся чаще всего в вакуолях. Они имеют вяжущий вкус и служат для защиты растений от поедания животными, а также обладают антисептическими свойствами. Таннины часто присутствуют в коре деревьев, незрелых плодах и листьях.
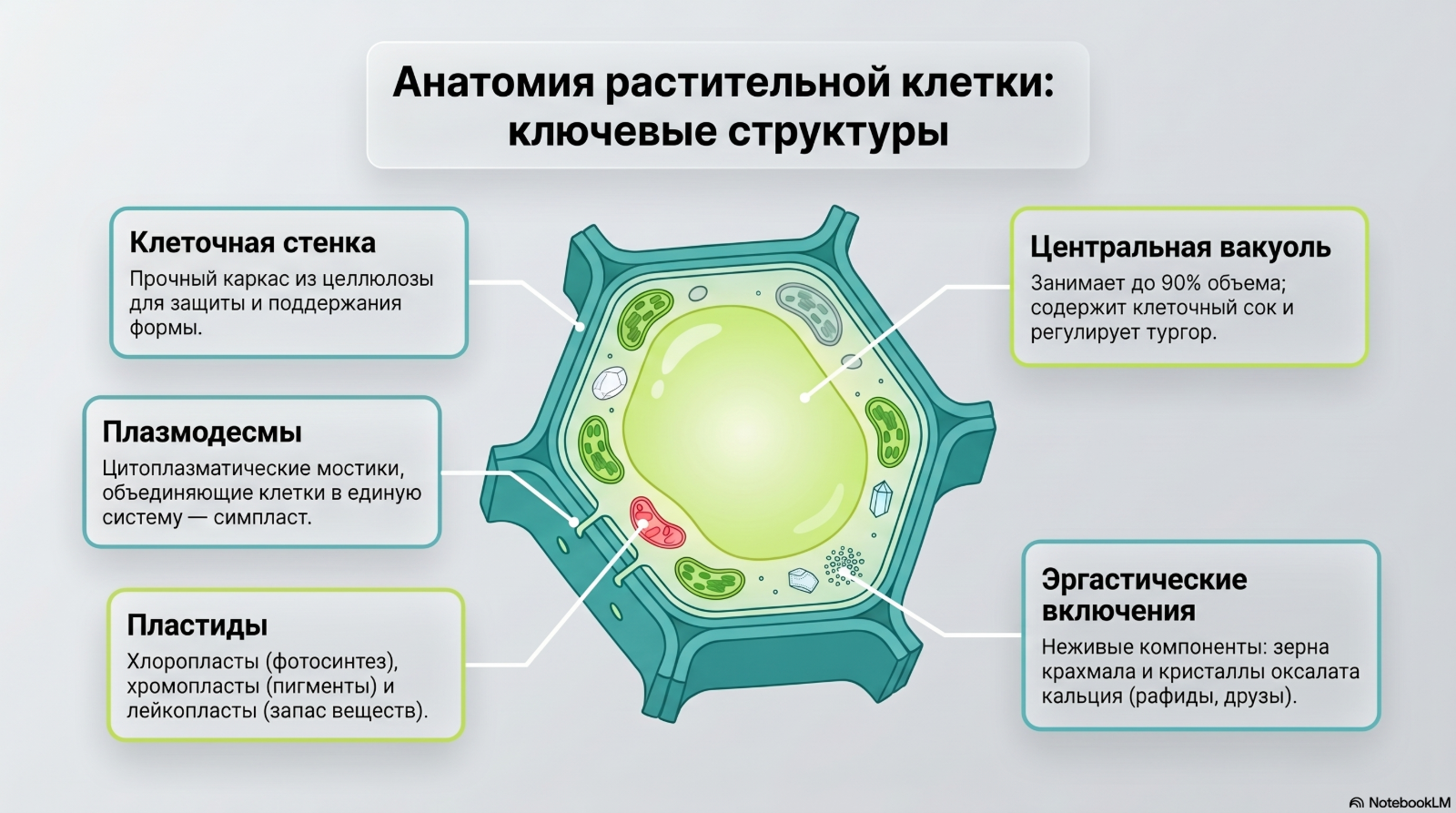
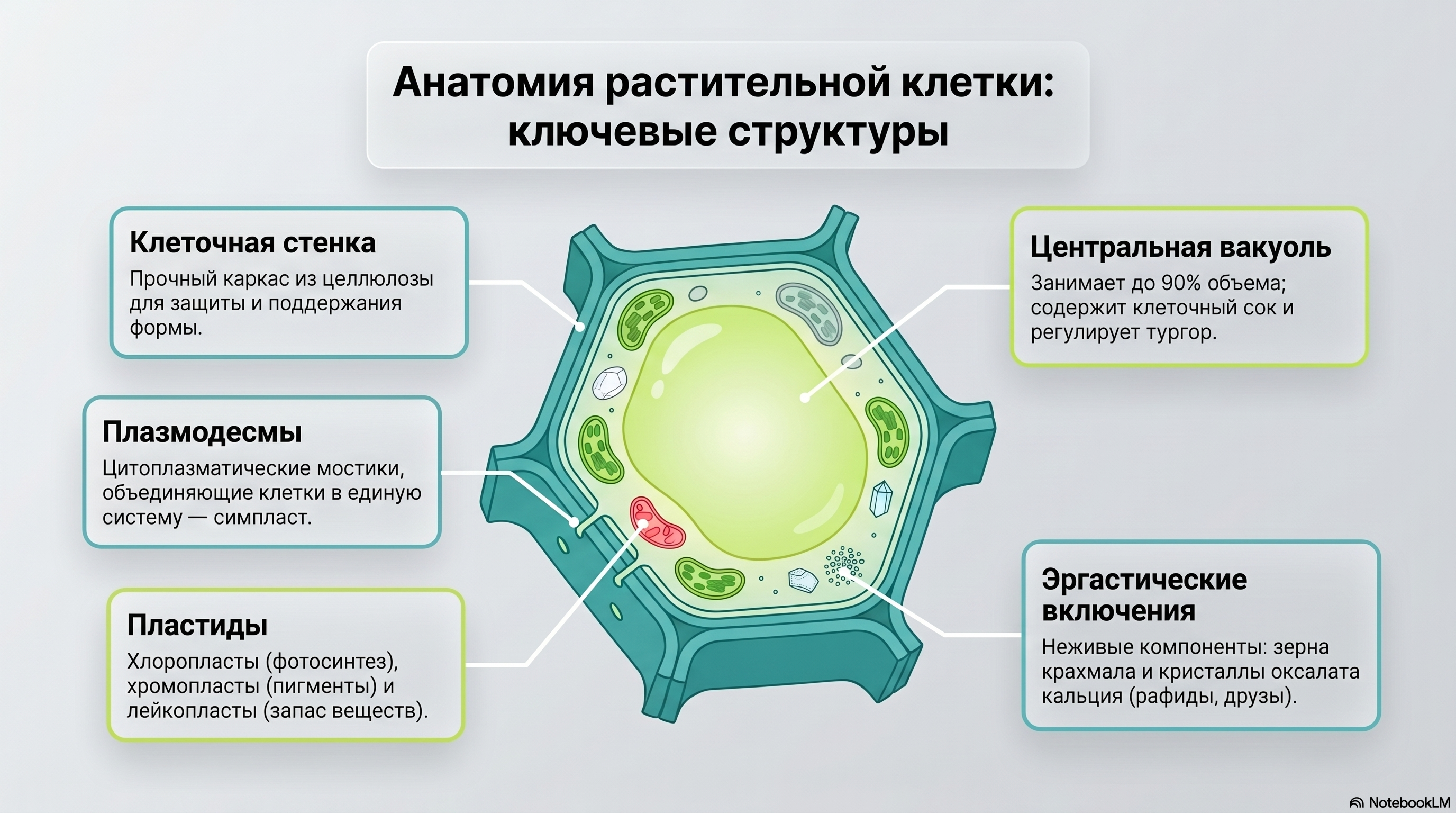 Рис. 7. Ключевые структуры растительной клетки
Рис. 7. Ключевые структуры растительной клетки