Щитовидная железа и её роль в эндокринной системе
Щитовидная железа — один из самых метаболически активных органов эндокринной системы, способный за считанные секунды реагировать на изменения йодного баланса и потребностей тканей. Её гормоны управляют скоростью обменных процессов почти в каждой клетке тела, влияют на рост, развитие нервной системы и работу сердца. Понимание того, как устроена железа, как она синтезирует гормоны и как эти сигналы достигают ядра клетки, позволяет увидеть целостную картину эндокринной регуляции.
Строение щитовидной железы и фолликулярный аппарат
Щитовидная железа состоит из множества замкнутых фолликулов диаметром от 100 до 300 мкм, заполненных коллоидом, который ограничен кубовидными эпителиальными клетками. Эти клетки выделяют секрет в просвет фолликулов, образуя запас гормональных предшественников. Коллоид представлен преимущественно тиреоглобулином — крупномолекулярным гликопротеином, в котором гормоны щитовидной железы находятся в связанном виде. Для поступления в кровоток они должны абсорбироваться через фолликулярный эпителий, что обеспечивает высвобождение активных гормонов.
Кровоснабжение железы характеризуется исключительной интенсивностью: минутный кровоток почти в пять раз превышает массу самой железы, что превосходит все другие ткани организма, кроме коры надпочечников. Такой приток крови гарантирует эффективный захват йодидов из плазмы и поддерживает непрерывный синтез гормонов.
Захват йода и йодное обеспечение организма
Базальная мембрана тироцитов обладает способностью активно захватывать йодид из крови и транспортировать его в клетки и фолликулы, функционируя как йодный насос (так называемая «йодная ловушка»). Этот процесс представляет первую стадию формирования гормонов — поступление йодидов в тироциты.
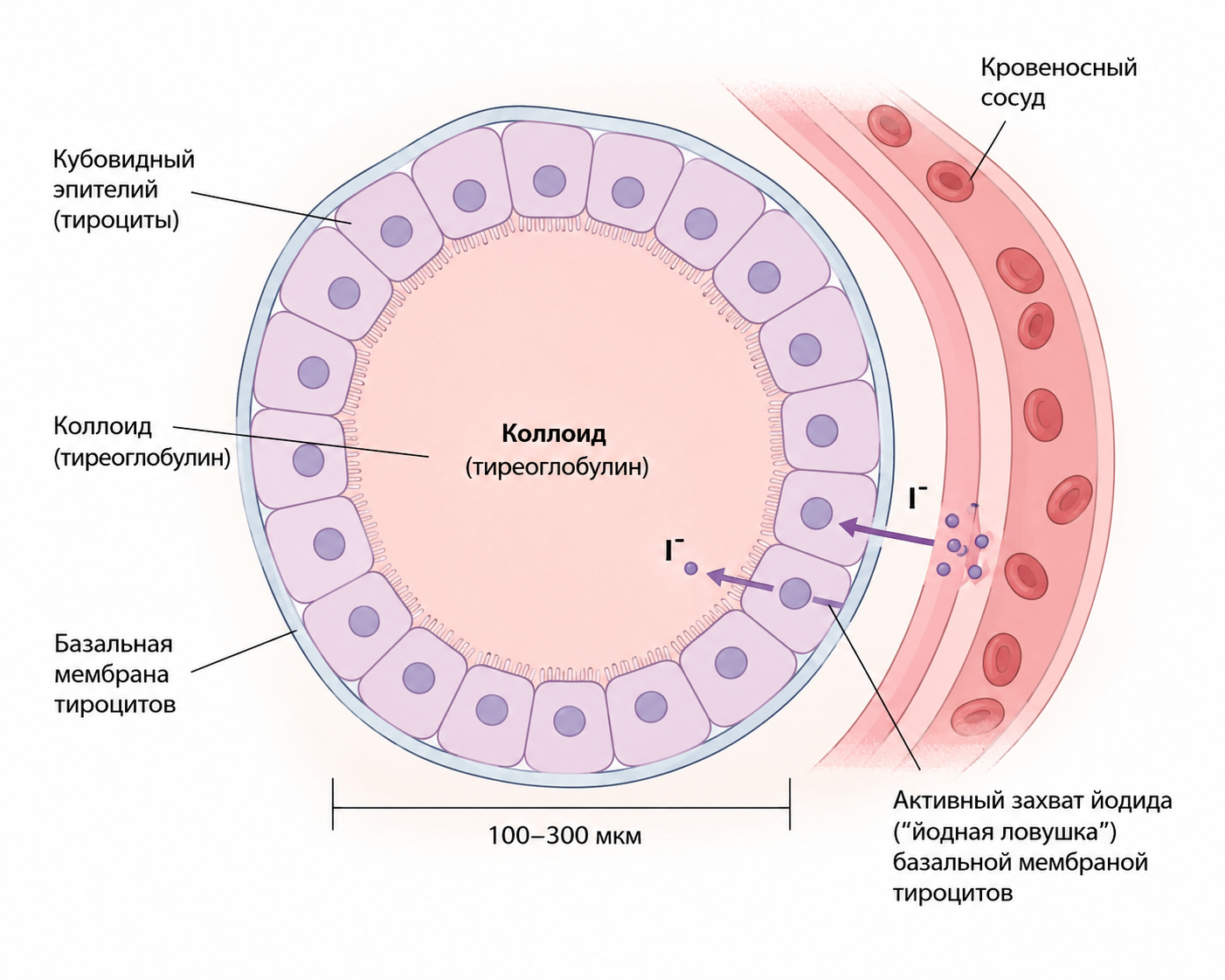 Рис. 1. Схема фолликула щитовидной железы. Видно коллоид с тиреоглобулином, кубовидный эпителий, активный захват йодида базальной мембраной тироцитов и кровеносные сосуды.
Рис. 1. Схема фолликула щитовидной железы. Видно коллоид с тиреоглобулином, кубовидный эпителий, активный захват йодида базальной мембраной тироцитов и кровеносные сосуды.
Для поддержания нормального уровня тироксина организму необходимо получать около 50 мг йода в год (примерно 1 мг в неделю). В качестве профилактической меры обычная столовая соль йодируется: к 100 000 частям хлорида натрия добавляют одну часть йодида натрия. Йодиды, поступающие с пищей, всасываются в желудочно-кишечном тракте аналогично хлоридам; большая часть выводится почками, а менее пятой части избирательно захватывается щитовидной железой для синтеза гормонов, что подчёркивает высокую селективность этого процесса.
При концентрации йодидов, превышающей физиологическую в 100 раз, отмечается подавление активности и уменьшение размеров железы. Этот эффект обусловлен снижением захвата йодидов, уменьшением продукции йодсодержащих гормонов и ингибированием эндоцитоза — механизма высвобождения гормонов из коллоида в кровоток. Благодаря этому свойству высокие дозы йодидов применяют за 2–3 недели до тиреоидэктомии, чтобы уменьшить объём железы и снизить кровопотерю при операции.
Гипертиреоидизм и болезнь Грейвса
При гипертиреоидизме (болезни Грейвса) щитовидная железа увеличивается в 2–3 раза за счёт гиперплазии и повышенной складчатости фолликулярного эпителия, а также роста числа клеток. Каждая тироцит секретирует гормон в несколько раз больше нормы, а общий уровень секреции гиперплазированной железы возрастает в 5–15 раз. Эти морфологические и функциональные изменения сходны с действием избытка тиреотропного гормона, однако у большинства больных концентрация ТТГ в плазме понижена или практически отсутствует. Вместо этого в крови обнаруживаются аутоантитела-иммуноглобулины, которые связываются с рецептором ТТГ и вызывают его продолжительную стимуляцию, поддерживая повышенную продукцию гормонов.
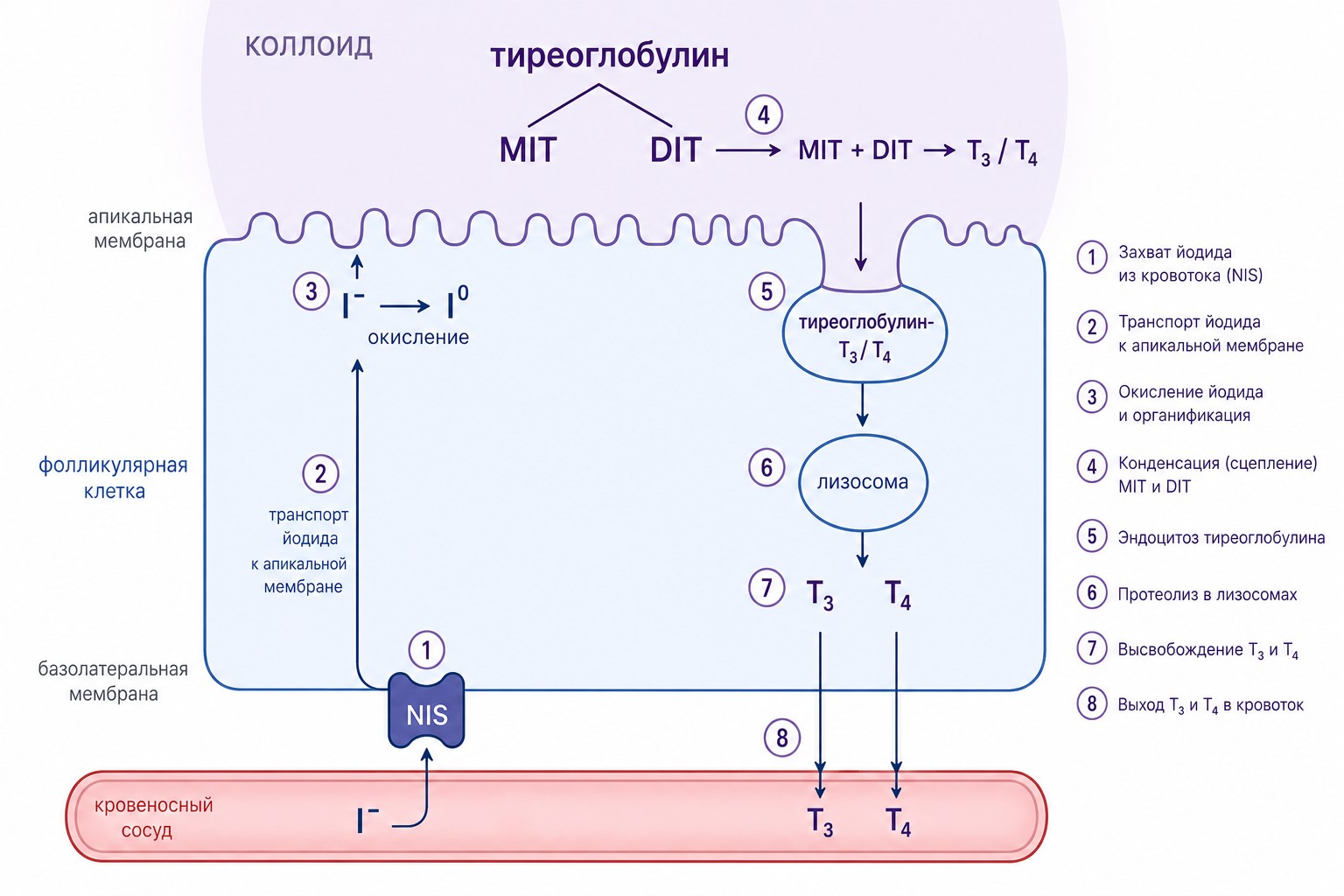
Синтез гормонов: от йодирования до органификации
Щитовидная железа захватывает иодид в виде иона I⁻, который поступает в фолликулярные клетки и окисляется ферментом пероксидазой, расположенным в апикальной мембране. Образующийся нейтральный йод (I⁰) непосредственно связывается с тирозиновыми остатками тиреоглобулина, запуская процесс органификации. Йодиназа ускоряет это йодирование, последовательно превращая тирозин в моноиодтирозин (MIT) и затем в дииодтирозин (DIT), сокращая время реакции с нескольких секунд до долей секунды и обеспечивая быструю доступность йодированных субстратов в коллоиде.
Органификация тиреоглобулина происходит в два этапа: сначала йодирование тирозиновых остатков до MIT и DIT, а затем соединение двух молекул DIT с образованием тироксина (Т₄) или одной молекулы MIT и одной DIT с образованием трийодтиронина (Т₃). Эти реакции протекают в коллоидном депо, где тиреоглобулин, синтезированный в аппарате Гольджи, высвобождается и взаимодействует с йодом. После формирования гормоны остаются ковалентно привязанными к тиреоглобулину до момента их высвобождения. При стимуляции клетки, например тиреотропным гормоном (ТТГ), происходит фагоцитоз коллоидных вакуолей в фолликулярную клетку, их слияние с лизосомами и протеолитическое расщепление тиреоглобулина, что освобождает свободные Т₄ и Т₃.
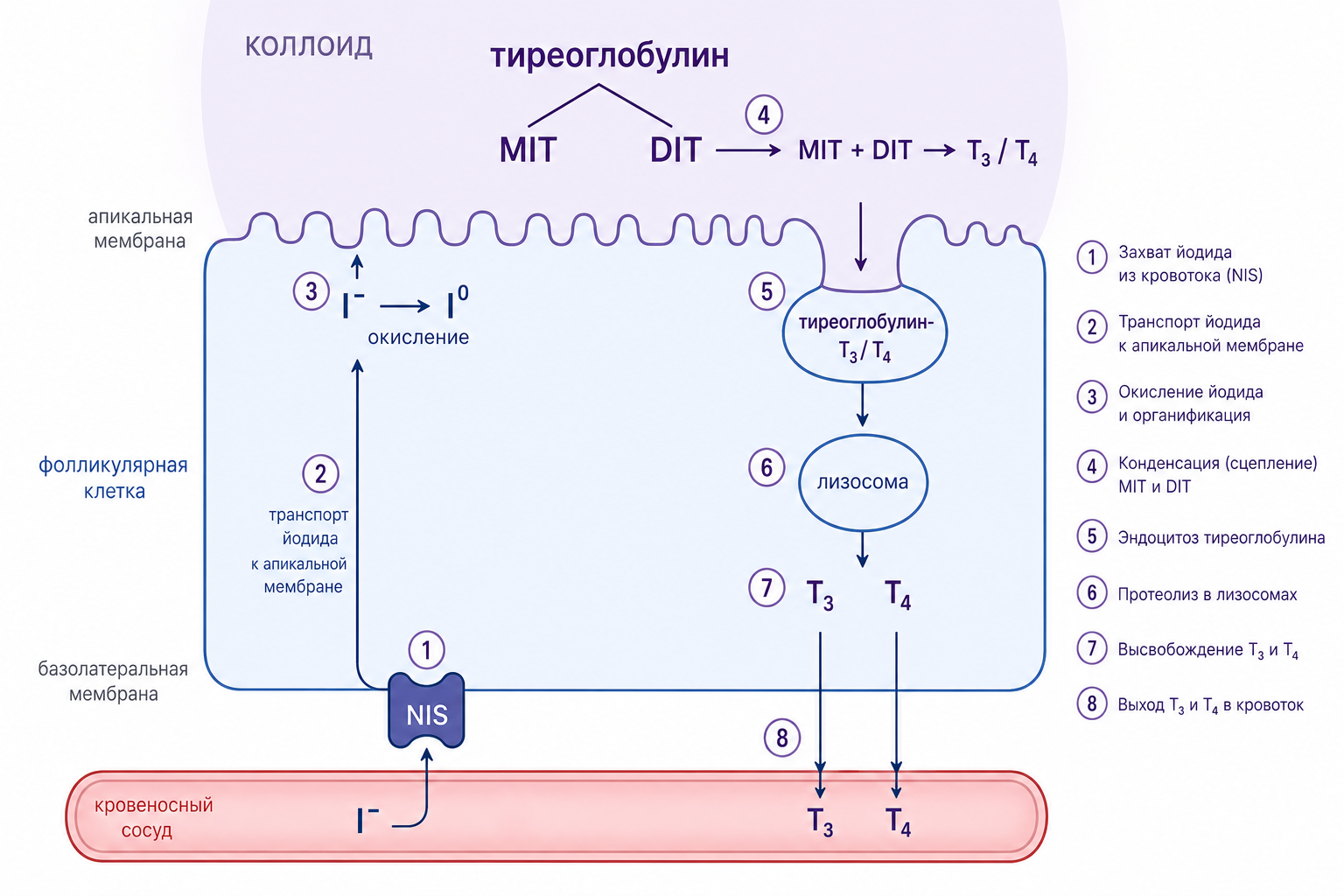 Рис. 2. Схема этапов синтеза и высвобождения гормонов щитовидной железы: йодирование тиреоглобулина, связывание MIT и DIT, формирование Т₄/Т₃, фагоцитоз коллоидного материала, лизосомальное расщепление, образование пиноцитозных везикул и экзоцитоз на апикальной мембране.
Рис. 2. Схема этапов синтеза и высвобождения гормонов щитовидной железы: йодирование тиреоглобулина, связывание MIT и DIT, формирование Т₄/Т₃, фагоцитоз коллоидного материала, лизосомальное расщепление, образование пиноцитозных везикул и экзоцитоз на апикальной мембране.
Высвобождение гормонов и транспорт в крови
Выделение гормонов сопровождается образованием пиноцитозных вакуолей, которые транспортируются к апикальной мембране, где происходит их экзоцитоз. Высокая концентрация пероксидазы на апикальной поверхности ускоряет процесс, обеспечивая окончательное окисление йода непосредственно при выходе тироксина и трийодтиронина в кровоток. Внутри фолликулярной клетки сохраняется запас до 30 молекул тироксина и лишь нескольких молекул трийодтиронина на одну молекулу тиреоглобулина, что позволяет железе поддерживать гормональный резерв в течение двух-трёх месяцев при отсутствии нового синтеза.
Нарушение функции пероксидазы (её блокада или отсутствие) полностью прекращает образование Т₄ и Т₃, поскольку без окисления йода процесс йодирования тирозина не происходит. После высвобождения гормоны связываются с транспортными белками плазмы (тироксин-связывающий глобулин, альбумин) и распределяются по тканям, где они регулируют метаболизм, рост и дифференцировку клеток.
Механизм действия на клетку: Т₄, Т₃ и ядерные рецепторы
Тироксин (Т₄) и трийодтиронин (Т₃) проникают в клетки-мишени посредством быстрой диффузии через их плазматическую мембрану. После входа в клетку большинство молекул Т₄ подвергаются дейодированию — отщеплению одного атома йода — что превращает их в активный Т₃. Таким образом, Т₄ функционирует в основном как прогормон, а биологически значимая часть Т₃ образуется непосредственно внутри целевых клеток, а не поступает в готовой форме из плазмы крови.
Трийодтиронин связывается с внутриклеточными рецепторами, которые представляют собой белковые комплексы, расположенные на хромосомах в ядре. Эти рецепторы образуют гетеродимер с ретиноид-X-рецептором и взаимодействуют с геномным фрагментом, реагирующим на тиреоидный гормон. При связывании Т₃ с таким ядерным рецептором происходит модуляция транскрипции: активируется или подавляется синтез определённых генов, что запускает производство белков-мишеней, отвечающих за клеточный ответ на гормон.
Главным эффектом тиреоидных гормонов является активация транскрипции сотен и более генов. Под их воздействием почти во всех тканях организма усиливается синтез ферментов, структурных и транспортных белков, а также других продуктов, что приводит к повышению общей функциональной активности клетки. Через эту генетическую программу реализуются многочисленные физиологические изменения: усиливаются сердечный выброс, кровоток, частота и сила сердечных сокращений, дыхание, потребление кислорода, активность Na⁺/K⁺-АТФ-азы, ускоряются захват глюкозы, глюконеогенез, гликогенолиз, липолиз, синтез белков и базовый обмен веществ.
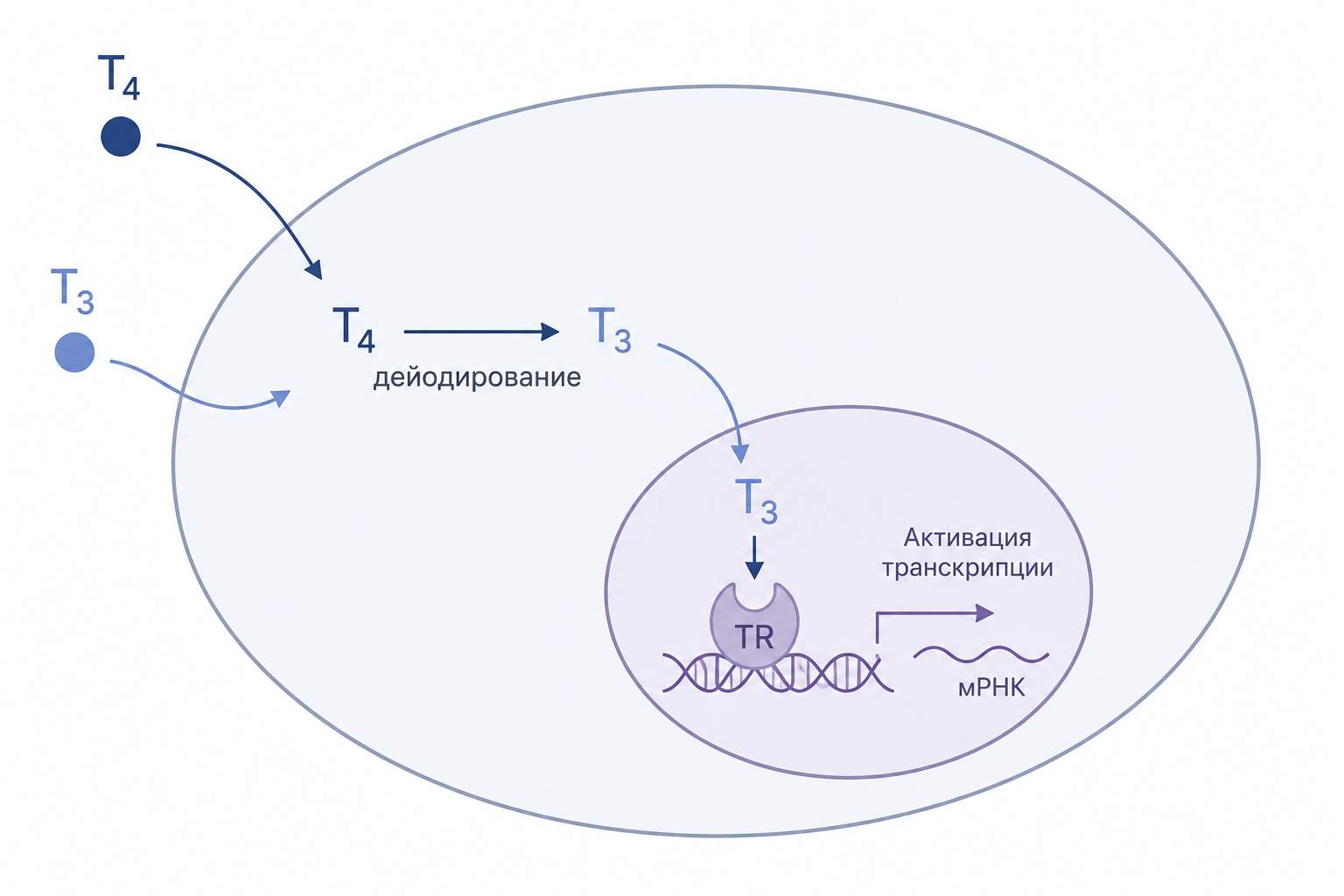 Рис. 3. Схема проникновения Т4 и Т3 в клетку. Показаны их внутриклеточное дейодирование, связывание Т3 с ядерным рецептором и последующая активация транскрипции генов.
Рис. 3. Схема проникновения Т4 и Т3 в клетку. Показаны их внутриклеточное дейодирование, связывание Т3 с ядерным рецептором и последующая активация транскрипции генов.
Однократное связывание гормонов с ядерными рецепторами обеспечивает длительный контроль экспрессии генов: эффект может сохраняться в течение нескольких дней и даже недель, поскольку активированный геномный аппарат остаётся «запрограммированным» на повышенную активность. Это объясняет длительный латентный период действия гормонов — после введения большой дозы тироксина заметное изменение скорости обменных процессов появляется лишь через 2–3 суток, а пик активности Т₄ достигает своего максимума через 10–12 суток. Трийодтиронин проявляет более быстрый эффект: латентный период составляет 6–12 часов, а максимум достигается уже через 2–3 суток.
Продолжительность действия гормонов определяется их распределением и связыванием с белками. В плазме крови гормоны в основном связываются с тироксин-связывающим глобулином, а в меньшей степени — с тироксин-связывающим преальбумином и альбумином. Такое связывание замедляет поступление Т₄ в ткани: для того чтобы половина циркулирующего Т₄ оказалась в тканях, требуется около 6 дней, тогда как Т₃, имеющий более низкое сродство к плазменным белкам, достигает половины своей тканевой фракции уже за сутки. После входа в клетки гормоны вновь связываются с внутриклеточными белками; Т₄ привязывается более прочно, чем Т₃, и хранится в клетках-мишенях, откуда высвобождается постепенно в течение нескольких недель. Период полувыведения тироксина составляет приблизительно 15 суток, а некоторые проявления его активности могут сохраняться от 6 недель до 2 месяцев.
Тиреоидные гормоны способны увеличить общий уровень основного обмена в тканях организма на 60–100 % выше нормы, активируя синтез и высвобождение метаболических гормонов в каждой клетке. При отсутствии их продукции основной обмен падает почти до 50 % от обычного уровня, что подтверждается данными, приведёнными на рисунке 76-6, где показана зависимость скорости базального метаболизма от концентрации гормонов.
Для поддержания очень высокого уровня базального обмена требуется одновременное повышение количества метаболических гормонов, синтезируемых щитовидной железой, поскольку их дефицит ограничивает способность тканей ускорять аэробные процессы и окислительное фосфорилирование.
Влияние на массу тела и энергетический обмен
Высокий уровень йодсодержащих тиреоидных гормонов почти всегда приводит к снижению массы тела, поскольку усиливается окислительный распад жиров и углеводов. При этом тиреоидные гормоны могут одновременно повышать аппетит, что частично компенсирует усиленный метаболизм и обусловливает неоднозначный эффект на массу тела.
Тиреоидные гормоны влияют на липидный обмен, снижая уровень общего холестерина в плазме: ускоряется утилизация липопротеинов низкой плотности и повышается экспрессия рецепторов LDL на клетках печени, что способствует более быстрому выведению холестерина из крови.
Они также усиливают метаболизм глюкозы, повышая чувствительность тканей к инсулину и ускоряя гликолиз. Одновременно стимулируется гликогенолиз в печени, что поддерживает высокий уровень глюкозы в крови и обеспечивает энергоснабжение ускоренного обмена.
Тиреоидные гормоны повышают концентрацию свободных жирных кислот в крови в периоды повышенной физической нагрузки за счёт ускоренного окисления жиров, что позволяет мышцам более эффективно использовать жирные кислоты для получения энергии. При гипертиреозе ускоряется мобилизация жиров из жировой ткани, однако это происходит не вследствие прямого действия гормонов на липолиз, а как следствие общего повышения энергетических расходов всех клеток, что приводит к повышенному расходу жиров как источника энергии. При длительном избытке тиреоидных гормонов может возникнуть состояние, схожее с кетогенным метаболизмом, поскольку ускоряется окисление жирных кислот и образование кетоновых тел; тем не менее основной механизм остаётся повышением общего энергетического обмена, а не прямой активацией липолиза.
Тиреоидные гормоны взаимодействуют с другими эндокринными регуляторами (инсулин, адреналин, кортикотропин), усиливая их метаболический эффект. Например, они повышают чувствительность к адреналину, что в совокупности активирует гормон-чувствительную триглицерид-липазу и ускоряет расщепление триглицеридов.
Влияние на сердечно-сосудистую и дыхательную системы
Тиреоидные гормоны усиливают кровоток и сердечный выброс, повышая метаболизм кислорода в тканях. Это достигается за счёт вазодилатации мелких сосудов, увеличения проницаемости капилляров к кислороду и ускорения удаления метаболитов, что приводит к вазодилатации и росту объёма крови, проходящего через кожу, и повышенной теплопередаче.
Увеличение сердечного выброса под воздействием тиреоидных гормонов может достигать 60 % от нормы, что сопровождается ростом частоты сердечных сокращений более существенно, чем ожидается лишь от увеличения объёма крови. Тиреоидные гормоны усиливают возбудимость миокарда, тем самым повышая частоту сокращений независимо от изменения нагрузки.
Тиреоидные гормоны повышают силу сокращений сердца, усиливая контрактильность миокардиальных волокон. Это связано с повышением уровня β-адренорецепторных сигналов и ускорением восстановления кальция в саркоплазме, что приводит к более эффективному сокращению сердца при каждом импульсе.
При гипотиреозе наблюдается снижение активности митохондриального дыхания, что приводит к уменьшению окисления как глюкозы, так и жирных кислот, тем самым замедляя общий метаболизм и способствуя накоплению липидов в тканях. Увеличение метаболизма под действием тиреоидных гормонов требует повышенного потребления кислорода и питательных субстратов, что приводит к росту потребности в кислороде и повышенной вентиляции лёгких, а также к увеличенной выработке тепла, что объясняет характерную потливость и чувство жара при гипертиреозе.
Регуляция функции: гипоталамо-гипофизарно-тиреоидная ось
Секреция тиреотропного гормна (ТТГ) передней долей гипофиза регулируется тиреотропин-рилизинг-гормоном (ТРГ), продуцируемым нейронами гипоталамуса. ТРГ поступает в портальную систему сосудов гипофиза, где стимулирует аденогипофиз к повышению продукции ТТГ. Таким образом формируется трёхуровневая ось регуляции: гипоталамус → гипофиз → щитовидная железа.
Тиреоидные гормоны (тироксин и трийодтиронин), попадая в кровь, действуют на переднюю долю гипофиза, подавляя секрецию ТТГ по механизму отрицательной обратной связи. Уже при увеличении уровня тиреоидных гормонов в 1,75 раза относительно нормы секреция ТТГ снижается практически до нуля, что свидетельствует о высокой чувствительности аденогипофиза к этим гормонам. Подавление ТТГ осуществляется преимущественно прямым действием тиреоидных гормонов на аденогипофиз; хотя часть эффекта может опосредоваться через снижение выброса ТРГ, большинство регуляторных проявлений обратной связи сохраняются даже при отделении аденогипофиза от гипоталамуса.
Благодаря этому механизму концентрация свободных фракций тиреоидных гормонов в плазме поддерживается почти постоянной, обеспечивая стабильность основного обмена и предотвращая как избыточную, так и недостаточную активность щитовидной железы.
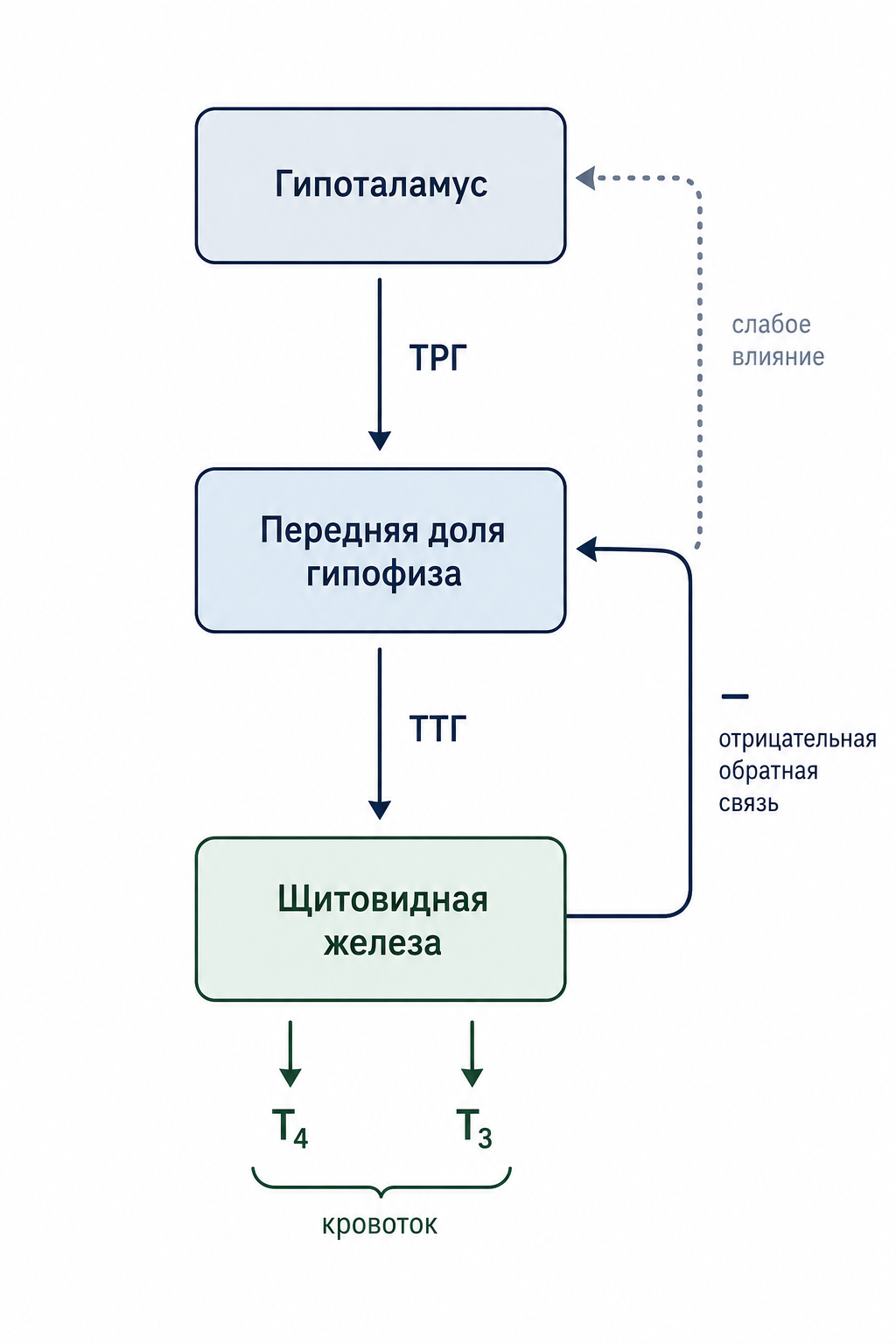 Рис. 4. Схема трёхуровневой оси регуляции функции щитовидной железы.
Рис. 4. Схема трёхуровневой оси регуляции функции щитовидной железы.
Переохлаждение организма или длительное пребывание в холоде активирует гипоталамус, усиливая секрецию ТРГ, что приводит к росту продукции ТТГ и, как следствие, повышению выброса тиреоидных гормонов — иногда более чем на 100 % относительно нормы. Этот процесс увеличивает основной обмен более чем на 50 %. Аналогично, эмоциональное возбуждение, тревожность и активация симпатического отдела автономной нервной системы стимулируют выброс ТРГ и ТТГ, повышая секрецию тиреоидных гормонов и теплопродукцию.
Эти влияния холода и эмоциональных факторов на функцию щитовидной железы исчезают при перерезке гипофизарного стебля, что указывает на то, что они реализуются исключительно через гипоталамус и требуют его связи с аденогипофизом. Для жителей арктических регионов характерно длительное повышение основного обмена на 15–20 % выше нормальных показателей, что отражает хроническую активацию гипоталамо-гипофизарной оси как адаптацию щитовидной железы к постоянному холодовому воздействию.
Роль в росте, развитии нервной системы и репродукции
Тироксин (Т₄), главный гормон щитовидной железы, ускоряет скорость почти всех химических реакций в клетках организма, тем самым повышая общий метаболизм и создавая энергетическую основу для роста и дифференциации тканей, включая ткани головного мозга в раннем детском возрасте. При отсутствии тироксина и трийодтиронина развитие нервной системы и рост мозга замедляются, что приводит к задержке умственного и физического развития у детей; именно поэтому гипотиреоз в детском возрасте считается одной из главных причин умственной отсталости.
Тироксин усиливает чувствительность гипофиза к гипоталамическому гормону, снижающему секрецию гормона роста (соматостатину), тем самым регулируя уровень гормона роста и обеспечивая согласованность роста костей, мышц и внутренней ткани.
Гормоны щитовидной железы положительно влияют на продукцию гонадотропных гормонов (ФСГ и ЛГ) в передней доле гипофиза, что происходит через усиление транскрипции генно-регуляторных элементов в клетках тиреотропных гормон-секретирующих нейронов гипоталамуса. Это опосредованное действие снабжает половые железы необходимыми стимулами для созревания и функции. При дефиците тиреоидных гормонов снижается выработка ФСГ и ЛГ, что приводит к задержке полового созревания, гипогонадизму и уменьшенному развитию вторичных половых признаков как у мальчиков, так и у девочек.
Тироксин взаимодействует с аденогипофизом, усиливая синтез гормона кортикотропина (АКТГ), что в свою очередь регулирует надпочечниковую продукцию кортикостероидов. Этим путём щитовидная железа участвует в поддержании водно-электролитного баланса и гомеостаза, влияя на условия, благоприятные для роста тканей. Гормон роста, секретируемый передней долей гипофиза, стимулирует рост большинства тканей тела, однако его анаболическое действие сильно усиливается при наличии адекватного уровня тироксина, поскольку тироксин повышает экспрессию рецепторов гормона роста на целевых клетках и усиливает сигнальные каскады (например, пути MAPK и PI3K/Akt).
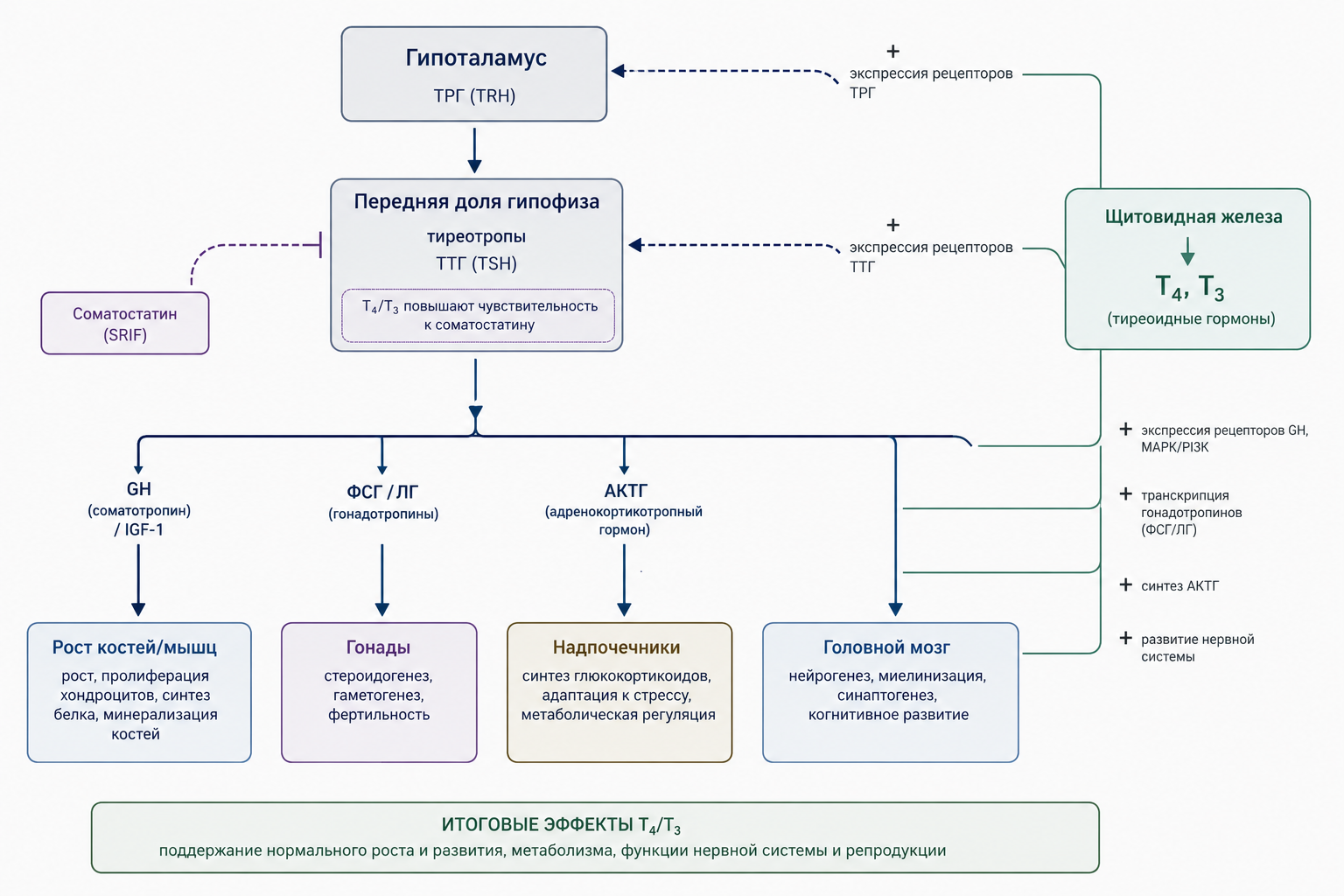 Рис. 5. Схема взаимодействия тиреоидных гормонов с гипоталамо-гипофизарной системой и их влияние на рост, метаболизм и репродукцию.
Рис. 5. Схема взаимодействия тиреоидных гормонов с гипоталамо-гипофизарной системой и их влияние на рост, метаболизм и репродукцию.
При гипертиреозе (избытке тироксина) наблюдается ускоренное развитие костей и рост организма, но одновременно может произойти преждевременное закрытие эпифизарных пластинок, что приводит к ограниченному росту в зрелом возрасте. При некоторых опухолях щитовидной железы, которые способны синтезировать тестостерон в количествах, превышающих норму более чем в 100 раз, происходит резкое увеличение мышечной массы и ускоренный рост костей у детей, однако избыточный андроген способствует раннему сращиванию зон роста, что в итоге уменьшает окончательный рост взрослого.
Тироксин также участвует в регуляции репродуктивных функций через прямое действие на гонадальные ткани: в яичниках он усиливает синтез стероидных гормонов, а в семенниках — поддерживает процесс сперматогенеза, что демонстрирует множественную роль щитовидной железы в репродуктивной системе. Нарушения в работе щитовидной железы могут влиять на функцию других эндокринных желёз, включая надпочечники, поджелудочную железу и половые железы, формируя комплексный эндокринный дисбаланс, который отражается на росте, развитии мозга и репродуктивных процессах.
Патологии щитовидной железы: коллоидный зоб и ферментные дефекты
Идиопатический нетоксический коллоидный зоб характеризуется значительным увеличением размеров щитовидной железы (в 10–20 раз), при котором наблюдается выраженный рост фолликулов, однако синтез тироксина и трийодтиронина в молекулах тиреоглобулина полностью отсутствует. Из-за невозможности продукции тиреоидных гормонов секреция ТТГ не подавляется, что приводит к прогрессивному увеличению фолликулов до огромных размеров. Точная причина развития идиопатического коллоидного зоба неизвестна, однако большинство исследователей указывают на связь с перенесённым в прошлом лёгким тиреоидитом.
Перенесённый тиреоидит, предположительно, вызывает небольшую гипофункцию щитовидной железы, что приводит к компенсаторной гиперпродукции ТТГ аденогипофизом и прогрессивному росту невоспалённых участков железы. Это объясняет очаговый характер увеличений: отдельные отделы щитовидной железы гиперплазированы, тогда как другие уменьшены в результате разрушающего влияния воспалительного процесса.
У некоторых пациентов с коллоидным зобом выявлены нарушения ферментных систем, необходимых для синтеза тиреоидных гормонов, включающие четыре основных дефекта: нарушение механизма йодного захвата (недостаточное поступление йода в тироциты), нарушение функции пероксидазной системы (снижение окисления йодидов), нарушение связывания йодированного тирозина в молекуле тиреоглобулина (делающее невозможным образование гормонов) и дефицит фермента дейодиназы, препятствующий восстановлению йода из йодированной формы тирозина, не объединившейся в молекулу тиреоидного гормона. Последний дефект ведёт к дефициту йода, поскольку приблизительно 2/3 йода возвращается в цикл через этот механизм.
Зобогенные вещества и фармакологическая блокада синтеза
Зобогенные вещества, содержащиеся в некоторых сортах репы и капусты, обладают активностью, подобной пропилтиоурацилу, и способны нарушать синтез тиреоидных гормонов. В результате их действия развивается гиперпродукция ТТГ, стимулирующая рост щитовидной железы и формирование зоба.
Тиоцианат действует как конкурентный ингибитор йодного захвата щитовидной железой: насос, транспортирующий ионы йода в тиреоидные клетки, также способен закачивать ионы тиоцианата, перхлората и нитратов. Поэтому введение тиоцианата в высокой концентрации вызывает конкурентное ингибирование транспорта йодидов в клетку (блокирование йодной «ловушки»). Снижение содержания йодидов в клетках не останавливает образование тиреоглобулина, но препятствует его йодированию и образованию йодсодержащих гормонов. Возникший дефицит тиреоидных гормонов приводит к увеличению секреции ТТГ аденогипофизом, что стимулирует рост железы, не продуцирующей достаточное количество гормонов, и вызывает формирование зоба.
Пропилтиоурацил (а также подобные ему соединения — метимазол и карбимазол) препятствует образованию тиреоидных гормонов из йодидов и тирозина путём блокады пероксидазы, необходимой для йодирования тирозина, а также блокады взаимодействия двух молекул йодированного тирозина, необходимого для образования тироксина и трийодтиронина. Пропилтиоурацил, как и тиоцианат, не препятствует образованию тиреоглобулина, однако отсутствие тироксина и трийодтиронина в тиреоглобулине вызывает мощную стимуляцию продукции ТТГ, приводящую к росту ткани щитовидной железы и формированию зоба.
Высокие концентрации йодидов (в 100 раз превышающие норму) способны уменьшать активность и размеры щитовидной железы, что представляет собой механизм, противоположный действию тиоцианата и пропилтиоурацила, которые вызывают зобогенный эффект.
Обратная связь и температурная адаптация
Увеличение содержания тиреоидных гормонов в крови снижает продукцию ТТГ передней долей гипофиза: если уровень тиреоидных гормонов превышает норму в 1,75 раза, продукция ТТГ падает практически до нуля. Практически все регуляторные проявления механизма обратной связи сохраняются даже при отделении аденогипофиза от гипоталамуса, что указывает на то, что ингибирование продукции ТТГ повышенным уровнем тиреоидных гормонов осуществляется их непосредственным влиянием на переднюю долю гипофиза.
Механизм обратной связи поддерживает концентрацию свободных тиреоидных гормонов в крови почти на постоянном уровне, что обеспечивает стабильность гормональной регуляции несмотря на колебания поступления йода и другие факторы среды.
Низкие температуры стимулируют выброс тиреоидных гормонов (иногда более чем на 100 % от нормы), что приводит к возрастанию основного обмена более чем на 50 %; у людей, оказавшихся в арктических регионах, основной обмен превышает нормальные показатели на 15–20 %. Эмоциональные реакции (общее возбуждение и тревожность), активирующие симпатический отдел вегетативной нервной системы, также могут вызывать гиперпродукцию ТТГ и повышать уровень обменных процессов и теплопродукции. Ни эмоциональные факторы, ни низкие температуры не являются эффективными на фоне перерезки гипофизарного стебля, что указывает на опосредованность этих реакций через гипоталамус.
Заключение
Щитовидная железа занимает центральное место в эндокринной системе, объединяя в себе функцию йодного накопителя, биохимическую фабрику по синтезу тироксина и трийодтиронина и эффекторное звено гипоталамо-гипофизарно-тиреоидной оси. От точности работы её фолликулярного аппарата зависят скорость основного обмена, терморегуляция, созревание нервной системы, рост тканей и состояние репродуктивной функции. Длительность действия тиреоидных гормонов, обусловленная их связыванием с плазменными белками и ядерными рецепторами, обеспечивает тканям устойчивый метаболический фон, а механизм отрицательной обратной связи удерживает концентрацию свободных гормонов в узких границах. Понимание этой системы — от захвата йодида до геномных эффектов Т₃ — позволяет видеть, как нарушения на любом этапе, будь то дефицит фермента, аутоиммунная стимуляция рецептора ТТГ или фармакологическая блокада синтеза, закономерно отражаются на обмене веществ, росте и развитии организма.