Строение листа
Листья — это настоящие биологические фабрики, обеспечивающие жизнь на нашей планете. Их поражающее воображение разнообразие форм и размеров скрывает глубоко продуманную внутреннюю архитектуру, где каждый тип ткани выполняет свою строгую функцию — от улавливания солнечного света до защиты от непогоды и транспорта веществ. Понимание анатомии листа позволяет увидеть, как именно растения адаптируются к окружающей среде, оптимизируя процессы фотосинтеза, газообмена и водного баланса.
Внешнее строение и морфология листовой пластинки
Листовая пластинка может быть расчленена на лопасти, доли или сегменты, что определяет её типологию: тройчатые, пальчатые и перистые листья различаются числом и расположением выростов. Отдельные формы расчленения часто сопровождаются специфическими очертаниями: у лировидных листьев преобладает крупная конечная доля, у прерывисто-перистых наблюдается чередование крупных и мелких долей, а струговидные характеризуются треугольными выростами. В сложных листьях, где пластинок несколько, каждая обычно имеет собственный черешочек (рахис), отходящий от общего черешка; отдельные пластинки называют листочками. Сложные листья бывают тройчато-сложными, пальчато-сложными (листочки в виде веера) и перистосложными (листочки вдоль рахиса в два ряда). Число листочков может быть парным или непарным; у некоторых растений встречаются дважды- и многократно-сложные листья.
Связь листовой пластинки со стеблем также важна для классификации. Если пластинка присоединяется к стеблю через черешок, такой лист называют черешковым; при отсутствии черешка и прямом соединении основанием — сидячим. В зависимости от того, насколько основания листа охватывают стебель, различают стеблеобъёмные (полное окружение) и полустеблеобъёмные (полулучевое охватывание) листья. В стеблеобъёмных листьях при сращении краёв основания образуется пронзённый лист; если основание образует продольную кайму, лист называют низбегающим, а стебель в этом случае характеризуется как крылатый. При разрастании основания в длину образуется влагалище, которое может частично или полностью обрастать стеблем в узле; степень срастания определяет полные и неполные узлы.
 Рис. 1. Связь листовой пластинки со стеблем.
Рис. 1. Связь листовой пластинки со стеблем.
Листовые пластины имеют дорзовентральное строение: адаксиальная (верхняя) сторона обращена к оси растения, а абаксиальная (нижняя) — от неё. Эти стороны могут отличаться по структуре и функциям. Размер листьев варьирует от нескольких миллиметров до нескольких метров; чем крупнее лист, тем их обычно меньше на растении. Крупные листовые пластины встречаются у древовидных папоротников и пальм (например, у корифы зонтичной до 8 м длиной и 6 м шириной, у рафии смолистой до 20 м × 12 м) и у водных растений, где пластинки виктории амазонской достигают 2 м в диаметре и выдерживают нагрузку 35–40 кг. Листовая пластинка обычно тонкая, зелёная и мягкая, но может принимать разнообразные текстурные формы: кожистые и жёсткие (падуб, самшит), пленчатые сухие и полупрозрачные (эфедра, казуарина), а также сочные и мясистые (агава, молодила, эхеверия). Зелёный цвет обусловлен хлорофиллом, который порой маскируется другими пигментами, например, антоцианами, растворёнными в клеточном соке, придавая листьям пурпурно-красные или голубоватые оттенки, как у клёна усечённого, барбариса Зибольда, синеголовника и некоторых бегоний. Серебристо-белые пятна на листовых пластинках представляют собой участки, где клетки содержат бесцветные пластиды — лейкопласты вместо хлоропластов, либо заполнены множеством межклетников, содержащих воздух, который рассеивает солнечные лучи.
Листовая мозаика представляет собой чередование листьев разных размеров пластинок и черешков, обеспечивая более равномерное освещение листьев одной плоскости. У клёна и конского каштана крупные листы с длинными черешками соседствуют с мелкими, что оптимизирует использование светового потока.
Эпидерма и устьичный аппарат
Эпидерма листа появляется в результате дифференциации протодермы, отделяющейся от остальных клеток верхушки побега под конусом нарастания. В поперечном срезе клетки эпидермы обычно имеют таблитчатую форму, при этом наружные стенки заметно толще антиклинальных (боковых) и периклинальных (внутренних) стенок. К наружным стенкам прикреплена кутикулярная покрывка — кутикула, которая у растений с толстыми стенками (например, агава, кливия, лук) состоит из нескольких кутинизированных слоёв, чередующихся со слоями целлюлозы. Кутикула нередко проникает между боковыми стенками, образуя дополнительные защитные структуры, а её неоднородное отложение формирует видоспецифический кутикулярный рисунок — бугорчатый, морщинистый, гребневидный или ячеистый. Над кутикулой обычно образуется эпикутанарный воск, придающий листовой поверхности сизоватый оттенок. Восковые отложения могут принимать формы чешуек, хлопьев, гранул или стерженьков. Наружные стенки основных эпидермических клеток могут содержать поры; у некоторых видов (к примеру, ковыля) поры простираются до кутикулы, тогда как в других они закрыты и образуют слепые, цокольные проходы.
Эпидерма состоит из нескольких типов клеток. Основные эпидермические клетки образуют преимущественную массу ткани; у злаков и осок они подразделяются на три морфологические группы: длинные, короткие вставочные и крупные пузыревидные (грушевидные в поперечном срезе). Замыкающие клетки устьиц находятся в непосредственной близости к устьицам, а трихомы (волоски) могут как присутствовать, так и отсутствовать. Многоклеточные волоски обычно имеют головчатую структуру, состоящую из ножки и головки. Железистые волоски отличаются от кроющих тем, что выполняют секреторную функцию, выделяя эфирные масла. Разнообразие опушения (бархатистое, шелковистое, мохнатое, войлочное, шерстистое, щетинистое, реснитчатое) определяется строением и плотностью волосков; верхняя и нижняя стороны листовой пластинки часто различаются по выраженности опушения. Крапива демонстрирует особый тип — жгучие трихомы, где кремниевый стержень с тонкостенной головкой легко отламывается при контакте, освобождая жидкость, богатую гистамином и ацетилхолином; у некоторых тропических видов такие волоски способны вызывать ожоги, приводящие к летальному исходу.
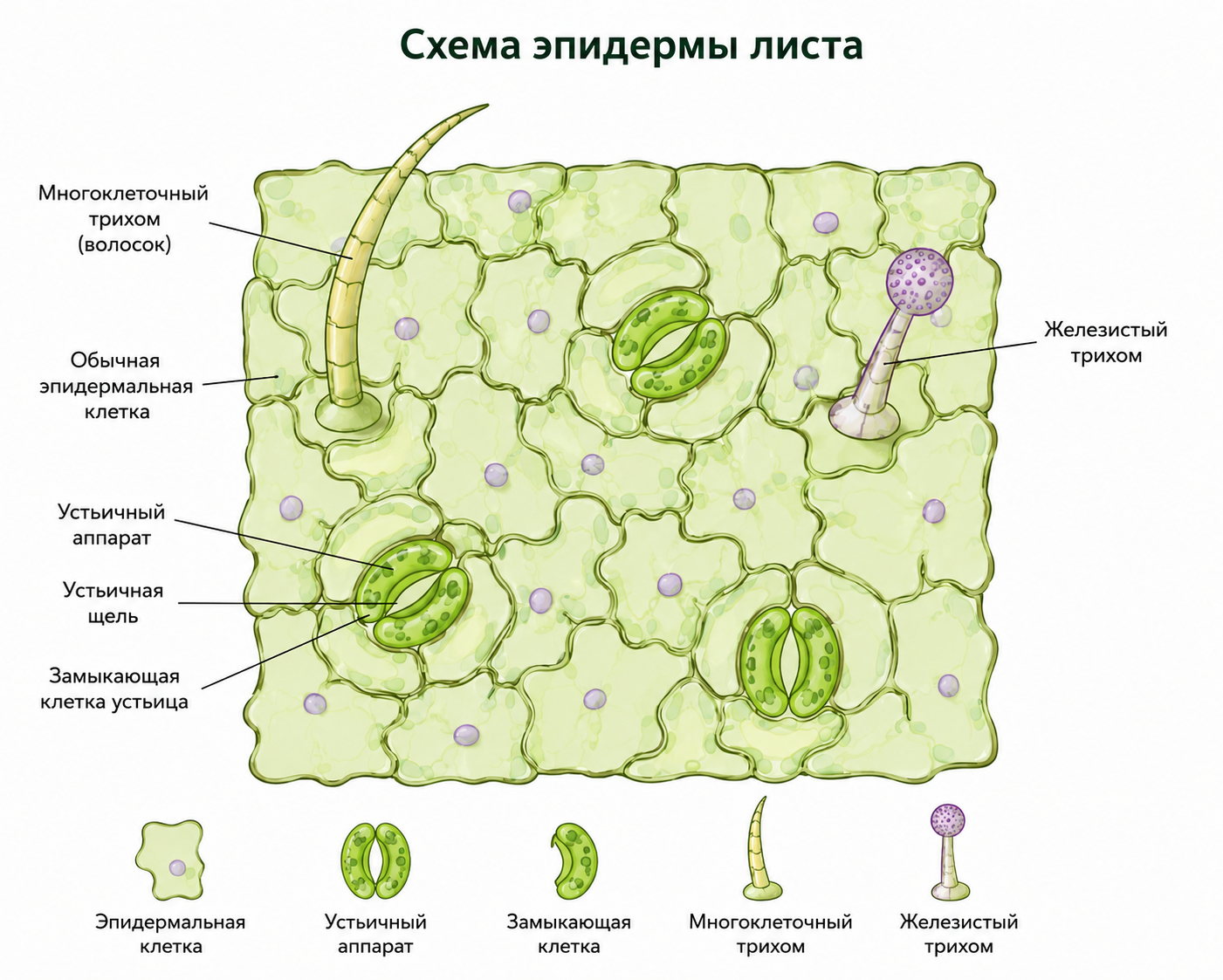 Рис. 2. Схема эпидермы листа
Рис. 2. Схема эпидермы листа
Устьица могут располагаться по-разному: на обеих сторонах листа (амфистоматические листья), только на нижней (гипостоматические) или только на верхней (эпистоматические). У плауна однолетнего наблюдается изменчивость: в основании побега листва эпистоматическая, в средней части — амфистоматическая или гипостоматическая, а на верхушке преобладают эпистоматические устьица. Плотность устьиц варьирует от 7 у традесканции до более чем 600 у лимона и паслена на 1 мм², отражая адаптацию к различным условиям среды; на неё существенно влияют влажность воздуха и интенсивность освещения. В узких листовых пластинках устьица образуют продольные ряды, а в широких — распределяются диффузно. Помимо устьиц, многие растения оснащены гидатодами — специализированными структурами, выделяющими капельно-жидкую воду.
Мезофилл и фотосинтез
У бузины чёрной в субэпидермальном слое мезофилла клетки обладают характерными складчатыми стенками, которые существенно увеличивают внутреннюю площадь оболочек. Такие клетки образуют односторонние складки и называются дланевидными. У большинства покрытосеменных растений мезофилл дифференцируется на два основных типа — столбчатый (палисадный) и губчатый. Столбчатый мезофилл обычно образует один-три слоя узко расположенных вертикально вытянутых клеток, ориентированных к свету, тогда как губчатый мезофилл состоит из более многочисленных слоёв с обильными межклетниками — воздухоносными полостями, оптимизирующими диффузию газов. Часто между этими типами располагается слой воронковидных (собирательных) клеток, стенки которых широки наружу, прилегая к столбчатому мезофиллу, и узки внутри, контактируя с губчатым. Собирательные клетки играют ключевую роль в отведении продуктов фотосинтеза из столбчатого мезофилла в губчатый.
В зависимости от расположения столбчатого мезофилла различают несколько типовых организаций листа. Бифациальный (дорзовентральный) тип характерен для двудольных: столбчатый мезофилл сосредоточен в верхней (дорзовой) части листа. Инвертированный бифациальный тип — обратная ситуация, когда столбчатый мезофилл находится в нижней (вентральной) части. Эквифациальный тип характеризуется одинаковой структурой мезофилла с обеих сторон листа, без ярко выраженного разделения.
Проводящая система листа
Внутренняя проводящая система листовой пластинки организована кольцевым расположением пучков в средней жилке; в нижних (абаксиальных) пучках ксилема обращена к верхней стороне пластинки, а в верхних (адаксиальных) — к нижней. В боковых пучках ксилема всегда находится в верхней части. Пучки, находящиеся в мезофилле, окружены однослойной паренхимной обкладкой, клетки которой вытянуты вдоль пучка и простираются до его окончаний, увеличивая площадь контактов. Боковые жилки различаются по составу: крупные похожи на пучки средней жилки, а с увеличением порядка ветвления количество проводящих тканей сокращается. Мелкие жилки состоят из нескольких трахеальных и ситовидных элементов и крупных сопровождающих клеток, при этом в ксилеме мелких пучков преобладают трахеиды со спиральными утолщениями. В флоэме мелких пучков сопровождающие клетки принимают фотосинтетические продукты из мезофилла через плазмодесмы, оболочки или промежуточный тип соединений, а затем водорастворимые сахара перемещаются в ситовидные элементы, где начинается нисходящий ток веществ.
Проводящие ткани листа и стебля представляют собой более сложную систему. Ксилема и флоэма образуют васкулярные меристемы — прокамбий и камбий. На ранних стадиях развития первичные ткани делятся на протоксилему и протофлоэму, а позже появляются метаксилема и метафлоэма. Первичные проводящие ткани присутствуют у всех высших растений, тогда как вторичные характерны преимущественно для голосеменных и двудольных. В коллатеральных и биколлатеральных пучках первичные ксилема и флоэма разделены вторичными проводящими тканями; между внешней ксилемой и второй флоэмой находится камбийная прослойка. Пучки с ней называют открытыми, в отличие от закрытых, где камбий отсутствует. У однодольных пучки часто располагаются диффузно без чёткого порядка. В стеблях спаржи ксилема приобретает V-образные очертания, а у рогоза ксилема и флоэма образуют вытянутые тангенциальные группы; метаксилема у рогоза состоит из двух широкопросветных сосудов, а у некоторых пальм — из единственного широкопросветного трахеидного сосуда. Над флоэмой часто развивается слой протофлоэмных волокон; у сусака и традесканции протоксилема разрушается рано, образуя водоносный канал. У однодольных встречаются концентрические амфивазальные пучки (например, в корневище ландыша). Межпучковая паренхима у однодольных часто содержит идиобласты, которые у ароидных (аронник, колоказия) становятся млечниками. У водных растений стелярная паренхима дифференцируется в аэренхиму. Перицикл, выраженный в стеблях некоторых однодольных (купена, лук, тюльпан, спаржа), состоит из многослойной одревесневшей склеренхимы. Диффузное распределение проводящих пучков препятствует образованию камбиального цилиндра, что ограничивает у большинства однодольных способность к вторичному утолщению.
Механические ткани
Механическая система листовой пластинки защищает лист от разрывов под действием ветра, дождя и града. Колленхима обладает уникальным строением: слои пектина чередуются со слоями целлюлозы, создавая состав, который сочетает высокую гибкость и значительную прочность. Влага составляет 60–70 % массы этих оболочек, причём её максимальное скопление наблюдается в угловых утолщениях. Неравномерное утолщение стенок и отсутствие одревеснения делают колленхиму пластичной, позволяя регулировать осмотическое давление за счёт изменяющегося тургора. При полном тургоре оболочки, пронизанные тонкими водопроницаемыми участками, функционируют как поры; без тургора поддержка утрачивается. Прочность колленхимы на разрыв превышает прочность литого алюминия, а её модуль упругости сопоставим со свинцом. В старых органах колленхима может затвердеть и стать хрупкой.
В отличие от неё, склеренхима характеризуется равномерно утолщёнными, обычно одревесневшими оболочками и разрушенными протопластами, представляя собой мёртвую ткань. Склеренхима состоит из двух типовых клеток: волокон (длинных прозенхимных клеток с острыми концами и простыми щелевидными порами) и склереидов (разветвлённых или длинных паренхимных клеток с многослойными стенками). Механические ткани листа функционируют в двух режимах: как балки-колонки (сопротивление статическим нагрузкам силы тяжести) и как балки-пружины (поглощение динамических воздействий). Распределение механической ткани отражает две тенденции: центробежную (ткани ближе к периферии, у молодых органов и низкорослых растений) и центростремительную (концентрация к центру, у высоких растений с массивной кроной). В боковых жилках наблюдается градация механической структуры: крупные жилки сходны с пучками средней жилки, а по мере ветвления количество проводящих тканей сокращается, влияя на прочность участков листа.
Анатомические адаптации к условиям среды
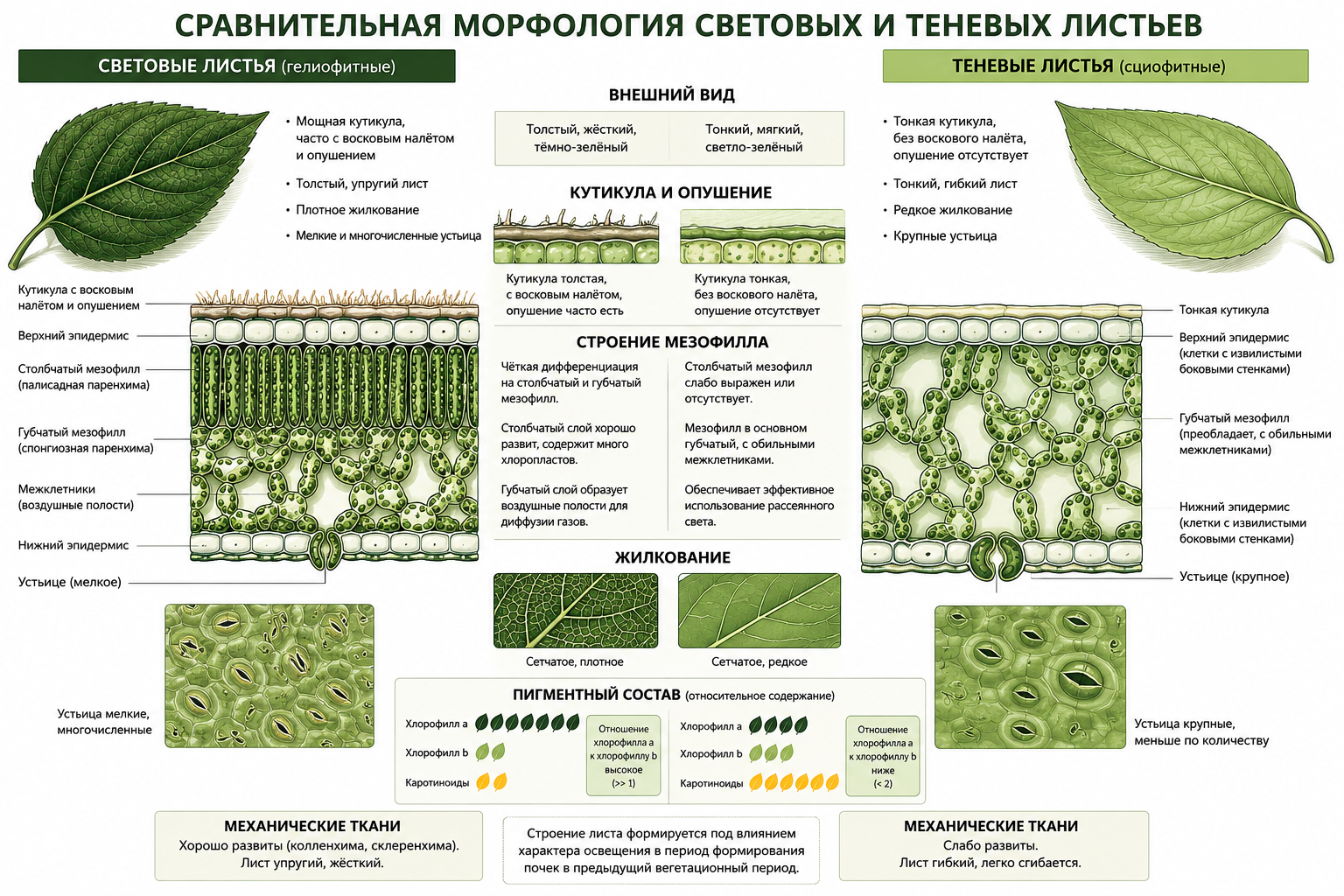
Структурное соотношение столбчатого и губчатого мезофилла сильно зависит от условий освещённости. Световые листья отличаются мощной кутикулой, часто покрытой восковым налётом и опушением, которые отражают избыточные солнечные лучи. Их эпидермальные клетки мельче, наружные стенки значительно утолщены, а боковые стенки ровные или слабо извилистые. Мезофилл световых листьев строго дифференцирован: столбчатый слой содержит большое количество хлоропластов, а губчатый образует воздушные полости для диффузии газов. Пигментный состав световых листьев отличается тем, что содержание хлорофилла а значительно превышает содержание хлорофилла в. Сетчатое жилкование у них более плотное, а устьица мелкие и многочисленные. Механические ткани световых листьев хорошо развиты, придавая листу упругость.
Теневые листья имеют тонкую кутикулу без воскового покрытия, а опушение обычно отсутствует. Наружные стенки их эпидермальных клеток менее утолщены, а боковые стенки сильно извилисты, что повышает площадь поверхности для газообмена. Теневые листья почти полностью состоят из губчатого мезофилла с обильными межклетниками. Отношение хлорофилла а к хлорофиллу в у них существенно меньше, а количество каротиноидов выше, что улучшает использование рассеянного света и защищает фотосистемы. Сеть жилкования развита слабее, устьица крупнее, а механических тканей гораздо меньше, что делает лист гибким. Строение листа формируется под влиянием характера освещения во время формирования почек в предыдущий вегетационный период.
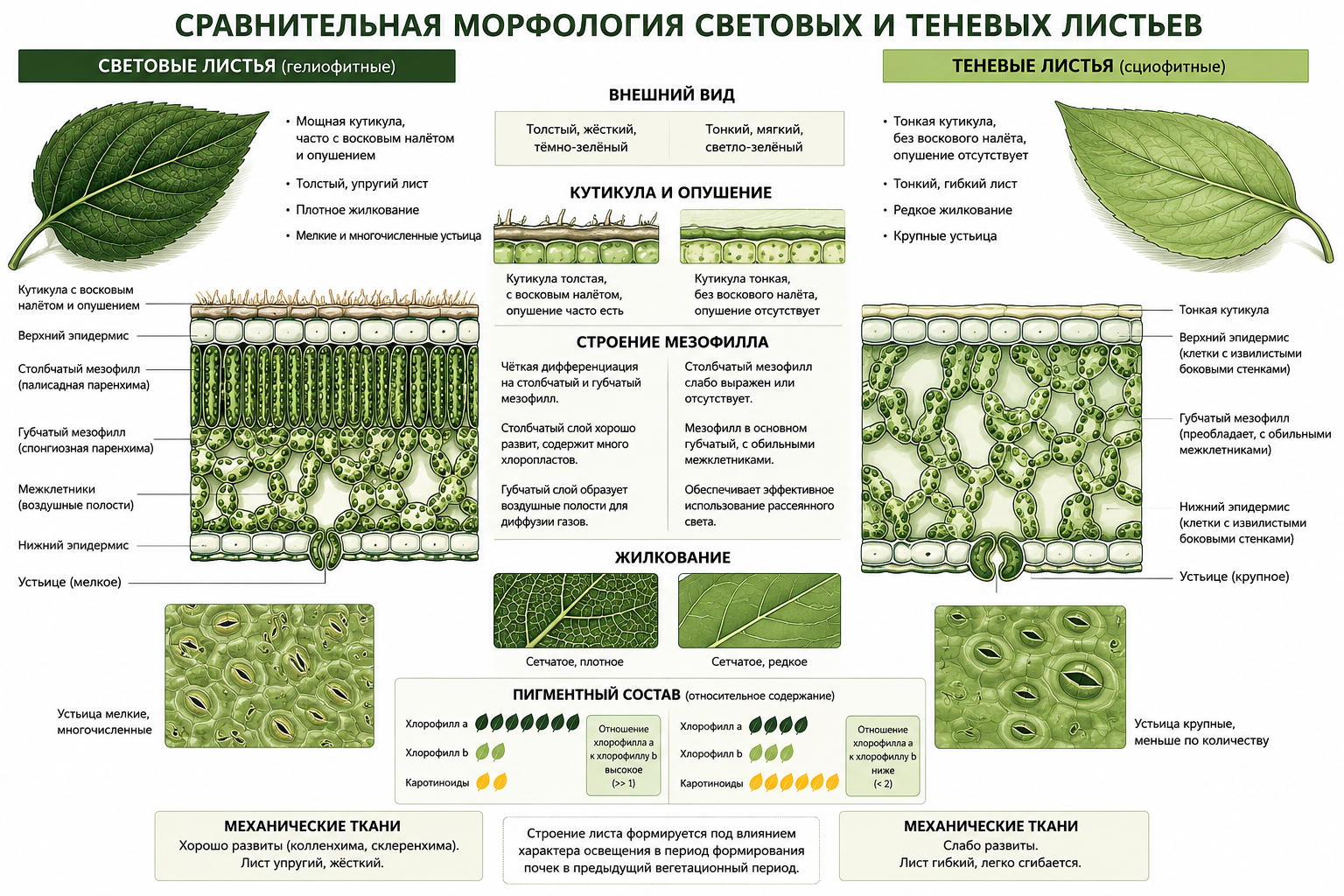 Рис. 3. Сравнение темновых и световых листьев: инфографика
Рис. 3. Сравнение темновых и световых листьев: инфографика
При постоянных односторонних ветрах крона деревьев и кустарников приобретает флаговую, однобокую форму. В арктических и высокогорных районах «снежная коррозия» стирает ветвления, формируя плоские и низкие кроны. В жарких условиях некоторые виды, такие как фикус, развивают трёхслойный эпидермис: внутренние прозрачные клетки запасают воду и задерживают тепловые лучи. В условиях сильного ветра растения укрепляют корневую систему (дуб, сибирская сосна, эвкалипт, секвойя), тогда как в условиях затенения светолюбивые виды образуют удлинённые междоузлия, а в полной темноте развивают этиолированные побеги.
Развитие и формирование листа
Листовой зачаток формируется как небольшая зачатка-конус на вершине растущего побега. У папоротников эта зачатка принимает форму трёхгранной пирамиды и делится параллельно её плоским сторонам, тогда как у двудольных инициальная клетка часто располагается субапикально. Рост поддерживается ритмичностью периклинальных и антиклинальных делений; у злаков (например, ржи) на верхушке листового примордия расположена группа инициальных клеток. Апикальный рост продолжается, пока длина не достигнет предела — от 0,8 до 3 мм у большинства растений и до 1,5 см у дудника. При этом происходит искривление зачатка из-за неравномерного разрастания абаксиальной и адаксиальной сторон. По достижении конечной длины начинается интеркалярный (вставочный) рост, сосредоточенный в основании будущей пластинки. У хвойных с игольчатыми листьями он ограничивается несколькими делениями, тогда как у однодольных продолжается значительно дольше.
Ключевым предшественником формирования пластинки является краевая (маргинальная) меристема, располагающаяся в виде двух симметричных валиков. Равномерные деления дают цельные листы; чередование активных и неактивных зон — расчленённые. У пальм листовые зачатки изначально простые, но в процессе развития образуют складки; при разрывах вдоль складок образуются «листочки», соединённые краевой полосой, которая позже отваливается. При длинных рахисах получаются перистые пальмовые листья, а при коротких — пальчатые. Весной, после опадения почечных чешуек, начинается внепочечное развитие: пластинки распрямляются, а между ними и основаниями листьев интеркалярно появляется черешок. Исследования И. Г. Серебрякова показали, что у черемухи площадь эпидермальных клеток удваивается, а площадь всей пластинки увеличивается в 1050 раз, что требует увеличения числа клеток более чем в 525 раз.
Листопад и отделительный слой
Листопад — характерный процесс для большинства растений умеренного климата, тогда как у голосеменных и субтропических или тропических покрытосеменных листы живут от трёх до пяти и более лет, опадая не одновременно. Существуют и альтернативные механизмы: отмирание надземных побегов у травянистых видов и постепенное высыхание с разрушением отмерших пластинок у папоротников и пальм. У африканской вельвичии функционируют лишь два листа, концы которых постепенно отмирают и расщепляются, а обновление обеспечивается интеркалярной меристемой в основании.
Перед опадом листья меняют окраску: у ольхи и сирени они опадают зелёными, у липы и берёзы желтеют из-за разрушения хлорофилла и обнажения каротиноидов, а у вишни, осины и клёна краснеют благодаря накоплению антоцианов. В этот период листовая ткань оттягивает синтезированные углеводы в зимующие органы, теряя азот и фосфор, но удерживая протеины, гемицеллюлозы, сахара и жиры. На основе листа образуется отделительный (разъединяющий) слой в основании черешка — плоская пластинка из мелких, тонкостенных клеток, насыщенных крахмалом, в которой почти отсутствуют механические ткани. У каштана, лоха и клёна этот слой формируется уже в начале весеннего развития, тогда как у большинства — за несколько недель или дней до листопада. В месте соединения отделительного слоя с проводящими пучками трахеальные элементы закупориваются тиллами (у некоторых — камедями и слизями), ситовидные пластинки закрываются мозолистыми телами из каллозы, а оболочки клеток ослизняются. После полного формирования отделительного слоя лист оказывается подвешенным лишь на тонких проводящих пучках, которые при малейшем напряжении легко разрываются.
Заключение
Анатомия листа представляет собой невероятно сложную и слаженную систему, где каждая ткань — от многослойной кутикулы до проводящих пучков — выполняет строго определённую функцию, обеспечивая выживание растения в условиях непрерывно меняющейся среды. Внешнее разнообразие листовых пластинок является лишь отражением их глубокой внутренней адаптации к уровню освещённости, влажности и механическим нагрузкам. Изучение строения листа, от закладки зачатка до формирования отделительного слоя при листопаде, раскрывает поразительную пластичность растительного организма, его способность эффективно утилизировать ресурсы и защищать себя, гарантируя продолжение жизненного цикла.